Proceedings
of The World Avocado Congress III, 1995 pp. 211- 216
RESPONSE
OF COTYLEDONS DETACHMENT OVER THE DEVELOPMENT OF NURSERY SEEDLINGS OF AVOCADO (Persea
americana Mill.)
Alejandro F. Barrientos-Priego, Manuel J. Morales-Nieto, and Gustavo Almaguer-Vargas
Departamento de Fitotecnia
Universidad Autónoma Chapingo
Chapingo C.P. 56230, Edo. de México, MÉXICO
Michal W. Borys
Universidad Popular Autónoma del Estado de Puebla
21 Sur 1103 cot. Santiago, Puebla
C.P. 62160, Edo. de Puebla. MÉXICO
Ma. Teresa Martinez-Damián
Fundación Salvador Sánchez Colin-CICTAMEX S.C
Ignacio Zaragoza No 6, Coatepec Harinas C.P. 51700
Edo. de México. MÉXICO
Facundo Barrientos Pérez
Centro de Fruticultura
Colegio de Postgraduados
Montecillo C.P. 56230,
Edo.de México MÉXlCO
Abstract
When cotyledons detachment
was practiced in avocado seedlings of 1-2 and 6-8 cm they did not survive more
than 28 and 35 days, respectively, because during these stages of development
the seedling depends totally on the cotyledon reserve. On the other experiments
when cotyledons removing was done on seedlings of more height (13-16, 20-25 and
30-35 cm), they survived more that the evaluation period, although a unequal
development was found for stem diameter, stem length and number of leaves. The
estimated number of leaves that assures the change of the function paper of the
cotyledons on the supply of reserves for development of the seedling is of 6 to
8 leaves.
1. Introduction
The propagation in Mexico of
the avocado (Persea americana Mill.) is carried out mainly by means of
sexual reproduction to obtain seedlings for subsequent grafting with selected
cultivars. The propagation is sometimes delayed by the lack of uniformity of
seed germination. This period of dormancy, being understood as such to the
endogenous causes that hinder the germination of the seed, even if the
environmental conditions will be optimum, cause that many plants reach the
conditions for grafting outside of the advisable time. Currently in the
commercial nursery plant production of avocado, there are several treatments to
obtain a rapid and uniform seed germination, among these; the removal of the
seed cover (Eggers, 1942), cuts on the apex of the cotyledons (Johnston and
Frolich, 1956), punctures (Martinez et al., 1969), among others. However, it
has not been evaluated the paper of the cotyledons on the initial development
of the plant. Exist preliminary evidences that the permanency of the cotyledons
favor the initial growth of the plantules of avocado (Flores-Meza, 1990a),
however, also were observed effects of not to favor it.
The objective of this
investigation was to evaluate the effect of the removal of the cotyledons in
different plant height, on the initial shoot growth of avocado.
2. Materials and methods
The present study was carried
out under glasshouse conditions at the Department of Fitotecnia of the
Universidad Autónoma Chapingo, Chapingo, State of Mexico.
The plant material that was
used in this work were "criollo" type avocado seeds of the Mexican
race (Persea americana var. drymifolia), obtained from a single tree at
Tenancingo, State of Mexico.
They were
completed a total of 5 experiments where 30 plants were used, on 15 of them the
cotyledons removal was practiced as variation factor and the other 15 plants
were used as controls. They were carried out Student's t tests with P--0.05, in
which the experimental unit was a plant with 15 replications.
To all the seeds the coats
were removed and treated with a solution of benomyl at the rate of I mg-liter
to prevent possible fungous infections. The embryos were sown directly in the
bags (15 x 30 cm) for all experiments, which were previously filled with
substrate formed by a soil and sand mixture in proportion of 3:1 and
disinfected with methyl bromide. Each experiment was begun when the plantules
of avocado reached a height within each interval, as follows: Exp. I.- I to 2
cm; Exp. 2.- 6 to 8 cm; Exp. 3.- 13 to 16 cm; Exp. 4.- 20 to 25 cm; and Exp.
5.- 30 to 35 cm.
In each experiment 30 plants
were selected, those which had reached the agreed height. The cotyledons
removal was accomplished with scalpel eliminating them completely and without
injuring the embryonic axis. Data of stem length and diameter, as well as
number of present leaves, was recorded weekly. Of these the increases
accumulated were analyzed statistically. The period of evaluation was of 4
months after the removal of the cotyledons.
3. Result
In the first two experiments,
after the removal of the cotyledons, the plants did not achieve to survive
beyond 28 and 35 days, while for the others three experiments, the plants
continued their development after 117 days.
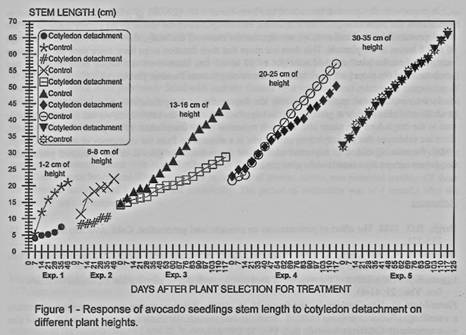
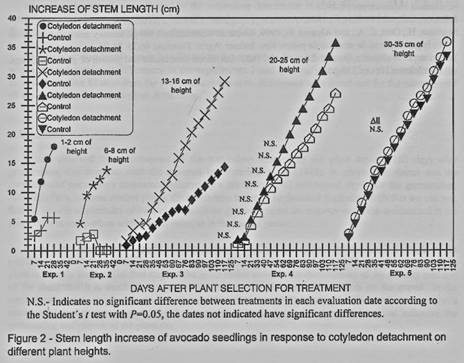
The effect of cotyledons
detachment over the stem length, in the first three experiments, shows a
reduction in the growth (figure I and 2). In experiment 4 until 62 days after
the cotyledons detachment there were no differences compared with the control
and for the experiment 5 they were not found differences.
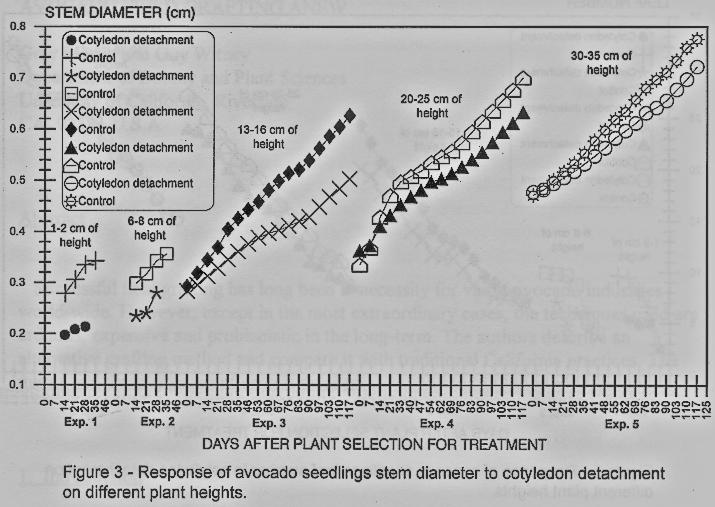
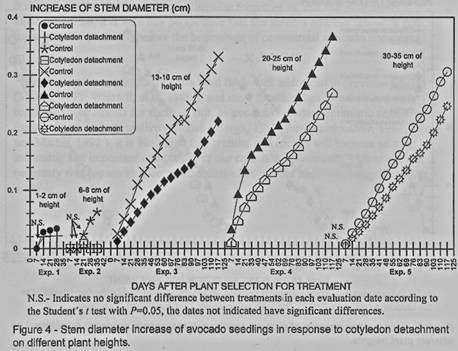
Stem diameter growth was
affected by the cotyledons detachment in all experiments where differences were
detected in almost all evaluation dates (figures 3 and 4), only in experiments
1, 2 and 5 some of the first dates there were no differences among the
treatment and the control.
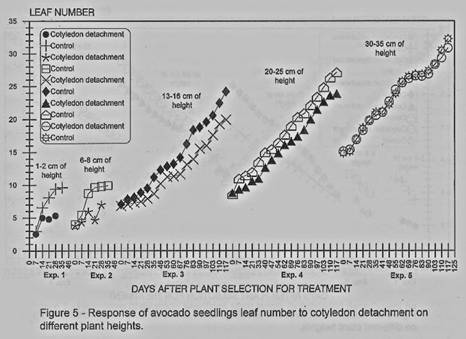
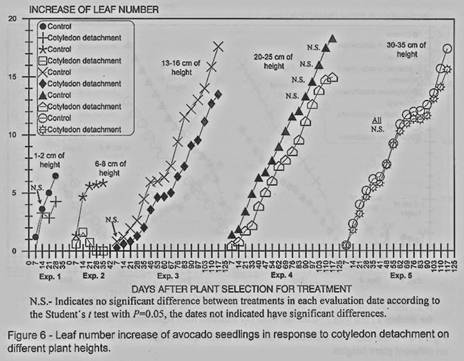
Leaf number was found also to
be affected by the cotyledons detachment in the first three experiments, with
some not significant differences in the experiments I and 3 in early stages
(figures 5 and 6). In the experiment 4 after 83 days from the cotyledons
removal no differences were found and for the experiment 5 all the evaluation
dates showed no differences.
4. Discussion
it was observed the
maintenance of the cotyledons adhered to the plant beyond 180 days after
germination, this contrasts with the reported by Wetzstein et al. (1983), in
which they found that the cotyledons of pecan were maintained adhered by a
minimum of 42 subsequent days after the germination. After this time, in some
control plants, the cotyledons began to be demoted gradually. With this we can
say, that the growth of plantules of avocado depends on greater time on the
reserves or on the synthesized in the cotyledons in comparison with other
species, as in the case of pecan.
Flores-Meza et al. (1990b)
found that the components of the size of the seed and residual weight of the cotyledons,
influence not only the germination, but also control the development of some
components of the plant in its early stages. The first stage of the germination
involves the development of the primary root and of the shoot that it is much
more slow. It is a heterothrofic stage that depends on the stored in the
cotyledons or of the synthetic activity. Blumenfeld and Gazit (1970) mention
that the seeds are not only a reservoir of growth regulators for the
development of the fruit, but also for citokinins that influence the
germination and growth of the plantules. In response to the question formulated
by Flores-Meza et al. (1990b), of which is the number of leaves that assures
the paper change of the cotyledons and acceptance of the leaves of the supply
function for the development of the stem and root, we can say, that the results
of our study, show that is required an average of 6 to 8 leaves in the
plantule. This does not mean that their function stops here, since until the
cotyledons removal when the plantules had a height of 30 to 35 cm, there were
not found differences for two components of the shoot part, more however,
concerning to stem diameter yet showed differences.
In the results it was
appreciated clearly the importance of the permanency of the cotyledons for the
production of plants ready to graft. Given the importance that have the
cotyledons, we can think that a cut or cuts to the cotyledons can cause also a
growth reduction in the components of the plant. For the moment, there are
evidences that the cotyledons cuts results in a stimulation from the emergency
of the shoot (Bergh, 1988). Because of this, it is important to evaluate the
response to growth of the produced plants after cotyledons cuts on the avocado
embryo or seed.
Reference
Bergh, B.O. 1988. The effect
of pretreatments on avocado seed germination. Calif Avocado Soc. Ybk. 72:
215-221.
Blumenfeld, A., and Gazit,
S., 1970. Citokinin activity in avocado seeds during fruit development. Plant
Physiol. 46:331-333.
Eggers, E. R., 1942. Effect of the removal of seed
coat o avocado seed germination. California Avocado
Soc. Ybk. 21: 41-43.
Flores-Meza, D.,
Vite-Cisneros, L., and Borys, M. W., 1990a. Relations among the componentes of
avocado seedlings (Persea americana Mill.). I. Seed size
components. II. Shoot size components. III. Root size components. California
Avocado Soc. Ybk. 74: 207-23 5.
Flores-Meza, D., Vite-Cisneros, L., and Borys, M. W., 1990b. Relaciones entre los cotiledones y otros componentes de plántulas de aguacate. CIC-FRUTICOLA-CICTAMEX Memoria 1990. Coatepec de Harinas. México. pp. 79-89.
Johnston, J. C., and Frolich, F., 1956. Avocado
propagation. California Avocado Soc. Ybk. 40: 89-98.
Martinez, H., Oste, C. A., and Alvarez, S., 1969.
Efecto de traurnatismos, temperaturas y sustancias quirnicas
en la germinación de semillas de paltos. Rev.
Industr. Agric. Tucuman 46(2): 81-96
Wetzstein, H. Y., Sparks, D.,
and Lang, G. A., 1983. Cotyledon detachment and growth of pecan seedlings.
HortScience 18(13):331-333.