Proc. of
Effect of Atmosphere Modification on Internal Physiological Browning of 'Fuerte' Avocados
A.B. Truter and F.J. Calitz
Department of Agricultural Development, FFTRI, Stellenbosch 7600,
J.G.M. Cutting
Department of Horticultural Science,
J.P. Bower
Outspan Citrus Centre, Nelspruit
1200,
Abstract. 'Fuerte' avocados were stored under controlled atmosphere (CA), modified atmosphere (CO2 shock) and regular atmosphere (RA) conditions for 4 weeks. Prior to, during and after storage, fruit samples were analyzed for PPO activity and ABA-content. Both PPO activity and ABA content were lowest in CO2 shock-treated fruit which indicated that these fruit may have been subjected to lower post harvest stress than fruit from the other treatments. PPO activity was highest in ripe CA-treated fruit, but these fruit had the lowest incidence of physiological browning. The highest incidence of browning was found in RA-treated fruit. CO2 shock can extend the storage life and maintain the quality of avocados because it decreases the potential for postharvest disorder development and is relatively easy and cost effective to apply.
South Africa is separated from its overseas markets by more than 9000 km and avocados exported to these markets can take up to 30 days to reach their destination, sometimes arriving in an advanced stage of maturity (Bower and Cutting, 1988). Lowering the shipping temperature to prevent this is not feasible due to the danger of chilling injury (Couey, 1982). In South Africa, it was shown that CA storage decreased the risk of chilling injury considerably (Truter and Eksteen, 1987). The optimum CA-conditions for avocados are 2% O2/10% CO2 at 5.5C (Hatton and Reeder, 1972; Truter and Eksteen, 1987).
Internal physiological disorders may also be induced by low temperature storage (Engelbrecht and Koster, 1986). Eaks (1976) found that a period of longer than 14 days at a temperature of 5.5C increases the risk of physiological disorders. In order to land South African avocados in Europe in an unripe state, exporters are forced to use temperatures around 5.5C during the shipping phase.
The most important physiological disorders of avocados are vascular abnormalities and mesocarp discoloration (Swarts, 1984). The potential for flesh browning of avocados is primarily a function of the browning enzyme PPO (Kahn, 1975). A further contributing substance is ABA which increases with maturity and ripening (Cutting et al., 1988; Cutting et al., 1990). Previous work by Spalding and Reeder (1974), Eksteen and Truter (1982, 1985) and Truter and Eksteen (1987) indicated that it may be possible to decrease the incidence of physiological disorders if a controlled or modified atmosphere is used during storage. However, the effect of these treatments on the biochemistry of the fruit has not been evaluated. Bower et al. (1990) studied the effect of CA, CO2 shock and RA storage on PRO activity in 'Fuerte' avocados. Both CA and CO2 shock-treated fruit showed a lower incidence of physiological disorders than the RA-treated fruit after 4 weeks storage. PPO activity was lowest in the CO2 shock-treated and highest in the CA-stored fruit. The results indicated that the fruit in CA storage for 4 weeks were in a state similar to that at harvest.
In an attempt to further elucidate the effect of controlled, modified and regular atmospheres on the internal quality of avocados and the role of PPO and ABA, 'Fuerte' avocados were subjected to these treatments and PPO activity and ABA-content were determined at weekly intervals over a 4 week storage period.
Materials and Methods
Fruit used in these trials were picked during May 1989 from irrigated 'Fuerte' trees at Westfalia Estate, Duivelskloof in the Northern Transvaal of South Africa. After normal pack-house treatment, the fruit were transported by road at approximately 6C to Stellenbosch, about 2000 km away. On arrival about 4 days after harvest, the following treatments were applied: CA storage at 2% O2 / 10% CO2 and CO2 shock using 25% CO2 with O2 decreasing to 1 % after 3 days. After treatment for 3 days, fruit of the latter treatment were stored in regular atmosphere. Fruit stored in RA from harvest served as controls. The total storage period at 5.5C from time of harvest was 32 days.
Five fruit from each treatment were randomly
selected for biochemical analysis on the day of arrival in Stellenbosch
(4 days) and subsequently at 7 day intervals for 32 days. The flesh (meso- and endocarp) of each fruit was cut into
approximately 10 mm cubes which were immediately frozen in liquid nitrogen and
freeze dried for later analysis. The specific activity of soluble PPO was
analyzed using the method of Bower and Van Lelyveld (1985) while
After 32 days, the remaining fruit were ripened at 20C until eating ripe and sampled again for biochemical analysis. The remaining ripe fruit were cut in half longitudinally and evaluated for the incidence of mesocarp discoloration and vascular abnormalities (Swarts, 1984). The experimental design was a one-way classification with three treatments (CA, CO2 shock and control) with 10 completely randomized replications (One experimental unit consisted of a carton of fruit). A working logit transformation (Snedecor and Cochran, 1980) was performed on the data before being submitted to analysis. Student's t-LSD was calculated to compare treatment means.
Results
PPO activity. Changes in PPO activity as influenced by treatment, storage time and maturity are shown in Figure 1. In the control fruit the PPO activity increased steadily from 4 days after harvest until the fruit reached the eating-ripe stage. In the CO2 shock-treated fruit, PPO activity remained almost unchanged during storage at 5.5C, but increased more rapidly during softening. In the CA-stored fruit, the PPO activity followed almost the same pattern as the other two treatments, except after 32 days when the differences between treatments were significantly larger. The lowest activity was in the CA-stored fruit, but during softening it increased sharply to reach the highest level of all treatments.
ABA-content. Prior to the application of the different treatments the ABA-content was high. Within 7 days of treatment (day 11) the ABA content had significantly decreased and remained more or less stable for the remainder of the storage period at 5.5C (Figure 2). During storage the control fruit generally had the highest ABA content, followed by the CA-stored and CO2 shock-treated fruit, respectively. After 32 days the ABA content in fruit of all treatments did not differ significantly. During softening, the ABA content of the CO2 shock-treated fruit remained almost at the same level as after 25 and 32 days, while those of the control and CA treatments increased significantly.
Internal fruit quality. The incidence of mesocarp discoloration was significantly higher in the ripe control fruit than in fruit of the other two treatments, but there were no significant differences between CO2 shock and CA-treated fruit (Table 1). Vascular abnormalities were also significantly higher in the ripe control fruit, compared with that of the other treatments. CA-treated fruit showed a significantly lower incidence of vascular abnormalities than CO2 shock-treated fruit.
Discussion
According to Kahn (1975) and Golan et al. (1977) there is a positive correlation between PRO activity and the tendency of avocado fruit to turn brown internally. Therefore, the browning potential of CA-stored fruit should have been the highest of all treatments (see Figure 1). However, when the avocados were examined internally, the opposite was found (Table 1).
According to Bower et al. (1990), CA storage
decreases the rate of postharvest physiological
changes in avocados to the extent that after 28 days, avocados under CA-storage
remained in a state similar to that at harvest. A freshly harvested 'Fuerte'
avocado in a non-stressed state has a specific soluble PPO activity of
approximately 5, which decreases rapidly during the early stages of ripening
(Bower et al., 1990; Cutting et al., 1990). In the
present study much lower PRO activity was found which is probably due to the
time lapse of 4 days before the first samples were analyzed.
The CO2 shock treatment resulted in the lowest PPO activity in eating-ripe fruit. However, the internal quality was also better than that of the control fruit. This confirms results of Truter and Eksteen (1987) and Bower et al., (1990). It therefore appears that the potential for physiological disorders was decreased by the CO2 shock treatment.
According to Cutting et al. (1990), PPO activity changed just before softening of avocados commences. This is probably the stage when browning takes place and the physiological disorders develop. During this process, PPO becomes inactivated, hence the lower activity where browning has occurred as opposed to CA where no browning has taken place (see Figure 1 and Table 1).
CO2 shock treatment holds promise as a storage technique, especially where CA storage facilities on ships are either not yet readily available or are available at high cost. A further advantage of this technique is that the fruit can be treated prior to export under RA-conditions, using simple, inexpensive equipment.
Literature Cited
Adato,
Bower, J.P. and J.G.M. Cutting. 1988. Avocado fruit development and ripening physiology. Hort. Rev. 10:229-272.
Bower, J.P., J.G.M. Cutting, and A.B. Truter. 1990. Container atmosphere, as influencing some physiological browning mechanisms in stored Fuerte avocados. Acta Hort. 269:315-321.
Bower, J.P. and L.J. van Lelyveld. 1985. The effect of stress history and container ventilation on avocado fruit polyphenol oxidase activity. J. Hort. Sci. 60:545-547.
Couey, H.M. 1982. Chilling injury of crops of tropical and subtropical origin. HortScience 17:162-165.
Cutting, J.G.M., J.P. Bower,
and B.N. Wolstenholme. 1988. Effect of harvest date and applied
Cutting, J.G.M., J.P. Bower,
B.N. Wolstenholme, and P.J. Hofman. 1990. Changes in
Cutting, J.G.M., P.J. Hofman, A.W. Lishman, and B.N. Wolstenholme. 1986. Abscisic acid, isopentyl adenine and isopentyl adenosine concentration in ripening 'Fuerte1 avocado fruit as determined by radioimmunoassay. Acta Hort. 179:793-797.
Eaks, I.L. 1976. Ripening, chilling injury and respiratory response of 'Hass' and 'Fuerte' avocado fruits at 20C following chilling. J. Am. Soc. Hort. Sci. 101:538-540.
Eksteen, GJ. and A.B. Truter. 1982. Beheerde en gemodifiseerde atmosfeer opberging van avokado's. S. A. Avocado Growers' Assn. Yrbk. 5:41-46.
Eksteen, GJ. and A.B. Truter. 1985. Effect of controlled and modified atmosphere storage on quality of eating ripe avocados. S. A. Avocado Growers' Assn. Yrbk. 8:78-80.
Engelbrecht, A.H.P. and
Golan, A., V. Kahn, and A.Y. Sodovski. 1977. Relationship between poly-phenols and browning in avocado mesocarp. Comparison between Fuerte and Lerman varieties. J. Agr. Food Chem. 25:1253-1260.
Hatton, T.T. and W.F. Reeder. 1972. Quality of 'Lulu' avocados stored in controlled atmospheres with or without ethylene. J. Am. Soc. Hort. Sci. 97:339-341.
Kahn, V. 1975. Polyphenol oxidase activity and browning by three avocado cultivars. J. Sci. Fd. Agric. 36:1319-1324.
Ludford, P.M. and L.L. Hillman. 1990. Abscisic acid content in chilled tomato fruit. HortScience 25:1265-1267.
Snedecor, G.W. and W.G. Cochran. 1980. Statistical methods.
7th ed.
Spalding, D.H. and W.F. Reeder. 1974. Low oxygen and high carbon dioxide controlled atmosphere storage for control of anthracnose and chilling injury of avocados. Phytopathology 65:458-468.
Swarts, D.H. 1984.
Truter, A.B. and GJ. Eksteen. 1987. Controlled and modified
atmospheres to extend storage life of avocados. S. A. Avocado Growers' Assn. Yrbk.
10:151-153.
Van Gorsel, H. and A.A. Kader. 1990. Phenolic metabolism and internal breakdown (chilling injury) of peach fruit. HortScience 25:1356.
|
Table
1. Incidence of mesocarp discoloration and vascular
abnormalities in 'Fuerte' avocados at the eating-ripe stage after 4 weeks
storage under three regimes. |
||||
|
Treatment |
Mesocarp discoloration |
Vascular abnormalities |
||
|
|
Logit |
% back transformation |
Logit |
% back transformation |
|
Controlled atmosphere (2%
O2/10% CO2) |
-2.323 |
8.9 az |
-2.073 |
11.2 a |
|
CO2 shock 25%
CO2 for 3 days |
-2.003 |
11.9 a |
-1.520 |
17.9 a |
|
Control |
-1.309 |
21.2 b |
-0.618 |
64.9 b |
|
LSD (P<0.05) |
0.6616 |
|
0.7923 |
|
|
z Mean
separation within columns by Student's t-LSD (P<0.05). Treatments with the
same letters do not differ significantly. |
||||
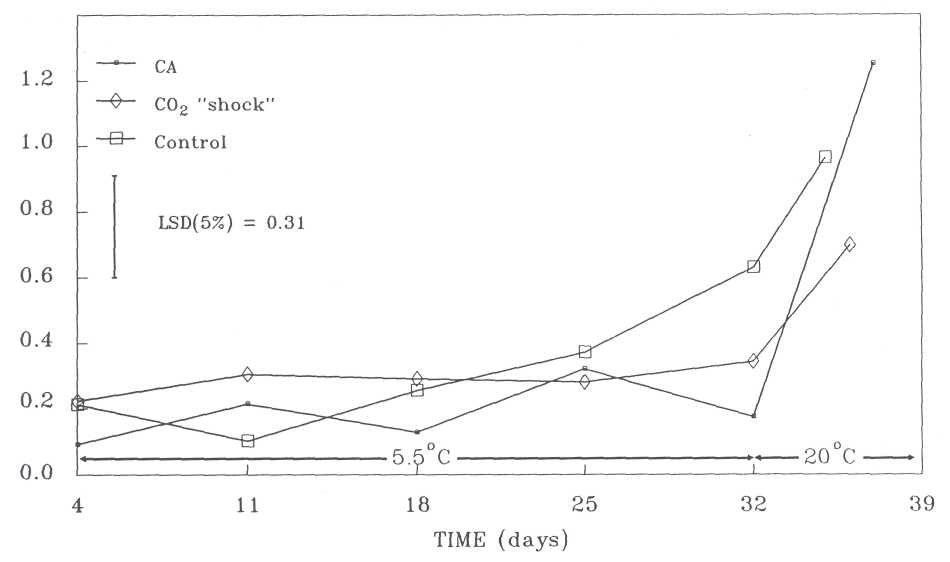
Fig.
1. PPO activity in 'Fuerte' avocados during storage at 5.5C and at 20C
(ripening stage).
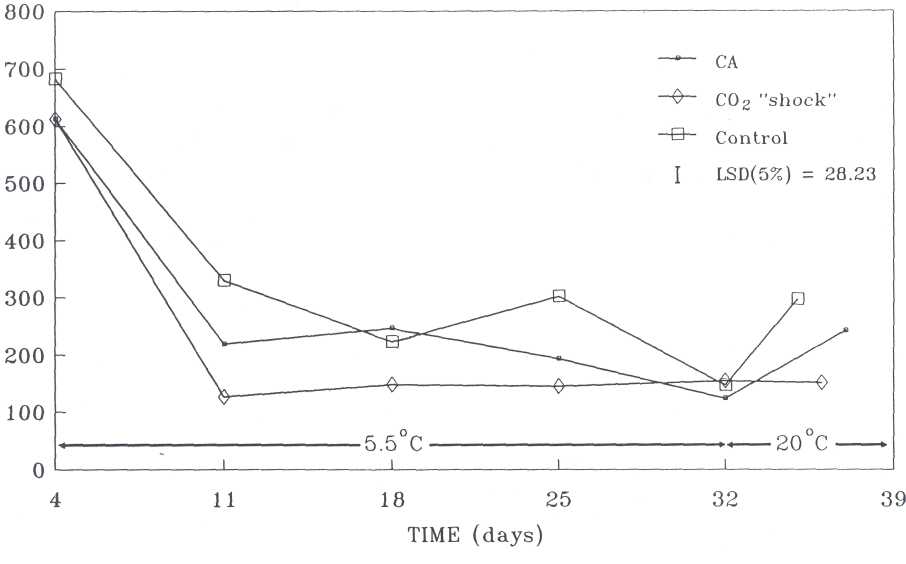
Fig. 2. ABA activity in 'Fuerte' avocados during storage at 5.5C and at 20C (ripening stage).