Proc. of Second World Avocado Congress 1992 pp. 237-240
Shoot Growth and Tree Architecture in a Range of Avocado Cultivars.
T. Grant
Thorp and Margaret Sedgley
Department of Horticulture, Viticulture and Oenology, Waite Agricultural Research Institute, University of Adelaide, Glen Osmond, SA 5064, Australia
Abstract. The objective of the research was to provide more information on avocado (Persea americana Mill.) tree architecture and how canopy design may be manipulated to optimize productivity. Avocado trees exhibit a monopodial, rhythmic pattern of growth, characteristic of the Rauh architectural model. Shoot modules are the smallest modular elements of this architectural model. They may form by prolepsis or syllepsis, with each type exhibiting a fixed, or predetermined, pattern of growth. The relative dominance of prolepsis and syllepsis is established by the interaction between apical dominance and acrotony. This interaction appears to be genetically determined and reflects cultivar differences in tree form.
Major advances in tree fruit production in crops such as apple have developed from a detailed knowledge of tree form and how this may be manipulated by tree training and pruning to optimize fruit yield and quality. This paper provides an architectural analysis of avocado tree growth for study of the effects of growth treatments and manipulation on productivity of avocado trees. The study of tree architecture, based upon architectural tree models, is a relatively new field of botanical science. It has introduced an element of precision to the "seemingly chaotic diversity" of tree form (Tomlinson, 1987). Twenty-three architectural tree models have been defined by Hall et al. (1978). Avocado is an example of the Rauh model, one of the most frequent among trees from temperate and tropical zones. It is characterized by monopodial, rhythmic growth. The main trunk forms branches morphogenetically identical to itself, and flowers are borne laterally with little effect on the vegetative shoot system.
Materials and Methods
Tree architecture of 'Sharwil', 'Gwen', 'Reed' and 'Hass' were analyzed using the following terminology. The terminology is based upon that used by Hall et al. (1978). In most tree species from tropical regions, growth is considered to be rhythmic. Shoot extension is periodic or seasonal, alternating with relatively short periods of rest. Each new period of rhythmic growth is called a growth flush and results in the formation of a growth module, hence rhythmic growth module (Fig. 1). An annual growth module is a section of annual growth and may include one or more rhythmic growth modules. The primary growth axis is the dominant growth axis in a growth module.
The smallest growth module is the shoot module. It is a "single unit of extension... (often) detectable between two successive groups of bud scale scars" (Hall, 1986). Shoot modules develop by either prolepsis or syllepsis. Proleptic shoots arise only after a period of dormancy as a resting bud. Hence, they have a bud scar ring at their base from the bud scales of the resting bud. Sylleptic shoots do not undergo a dormant period and have no bud scar ring. Sylleptic shoot growth is contemporaneous with extension of the parent axis. A branch is a complex of shoot modules formed over more than one annual growth flush. Major limbs are the largest growth modules. They are large branches which over one or more years have become major structural elements of the tree.
The control of axillary shoot formation in plants has been traditionally considered in terms of apical dominance. Strong apical dominance is the complete inhibition of lateral bud growth; weak apical dominance involves only partial inhibition. Apical dominance, however, only controls the growth of lateral buds on current season's shoots. Apical control in subsequent seasons, determining the release of subterminal resting buds, is termed acrotony. Champagnat (1978) described acrotony as the release of daughter axes that allows them to become dominant; and apical dominance as the suppressive effect of a parent bud on the daughter bud it produces.
This
research tests the hypothesis that differences in tree habit between avocado
cultivars are established by factors controlling the relative degrees of
prolepsis and syllepsis in rhythmic growth modules.
Results and Discussion
Strong genetic control over modular construction, that appeared to be species- rather than cultivar-specific, was indicated by the predetermined growth of avocado shoot modules. All shoot modules contained, on average, the same number of nodes with little variation, regardless of cultivar, season or location. A predetermined pattern of growth was also recorded on the orthotropic axis of annual growth modules, and on the primary growth axes of rhythmic growth modules. Similar patterns of predetermined growth have been recorded in trees from tropical (Barthelemy, 1986; Hall et al. 1978), and temperate (Lanner, 1976) regions.
Superimposed on this species-specific modular construction were important cultivar differences between the number of major limbs, their angle of growth, and the relative dominance of axillary shoot growth. A branch hierarchy was established during the first stages of shoot growth and appeared to become entrenched in later years. Thus, cultivar differences in tree habit were reflected in the pattern of axillary shoot formation on rhythmic growth modules.
A positive correlation was found in all avocado cultivars studied between apical dominance and acrotony such that strong apical dominance involved the inhibition of syllepsis, and strong acrotony involved the release of proleptic buds. Thus 'Sharwil' exhibited strong apical dominance with relatively few sylleptic shoots, but strong acrotonous growth with many major limbs and long and numerous proleptic shoots. In contrast, 'Gwen' and 'Reed' displayed weak apical dominance in that they had large numbers of sylleptic shoots, although these were relatively short. They also displayed weak acrotony, producing few major limbs, relatively short axillary shoots and few proleptic shoots. 'Hass' was intermediate between 'Sharwil' and 'Reed' and so were the relative influences between apical dominance and acrotony on axillary shoot growth. This information supports the hypothesis that cultivar differences in tree habit are established by factors controlling prolepsis and syllepsis in rhythmic growth modules.
Conclusions
What are the horticultural consequences of predetermined growth, and modular construction? Is there an ideal module design for optimum orchard productivity? More compact growth habit, e.g., 'Reed' compared with 'Hass', appeared to be linked with weak apical dominance and high syllepsis. Plant growth regulators and certain rootstocks may be useful to encourage this type of growth. Pruning, to rejuvenate modular construction on older trees, may improve average module productivity. Removal of vigorous proleptic shoots from young 'Sharwil' trees may encourage more compact modular growth with this cultivar. Analysis of modular construction provides a simple means of interpreting the effects of growth treatments and manipulations. It will also have universal application with other trees which exhibit rhythmic growth.
Literature Cited
Barthelemy, D. 1986. Establishment of modular growth in a tropical tree: Isertia coccinea Vahl. (Rubiaceae). Philos. Trans. R. Soc. Lond. 313:89-94.
Champagnat, P. 1978. Formation of the trunk in woody plants. In: P.B. Tomlinson and M.H. Zimmermann (eds.). Tropical trees as living systems. Cambridge University Press, pp. 401-422.
Hall, F. 1986. Modular growth in seed plants. Philos. Trans. R. Soc. Lond. 313:77-87.
Hall, F., R.A.A. Oldeman, and P.B. Tomlinson. 1978. Tropical trees and forests: an architectural analysis. Springer Verlag, Berlin, Heidelberg, New York.
Lanner, R.M. 1976. Patterns of shoot development in Pinus and their relationship to growth potential, p. 223-243. In: M.G.R. Cannell and F.T. Last (eds.). Tree Physiology and Yield Improvement. Academic Press, London, New York, San Francisco Tomlinson, P.B. 1987. Architecture of tropical plants. Ann. Rev. Ecol. Syst. 18:1-21.
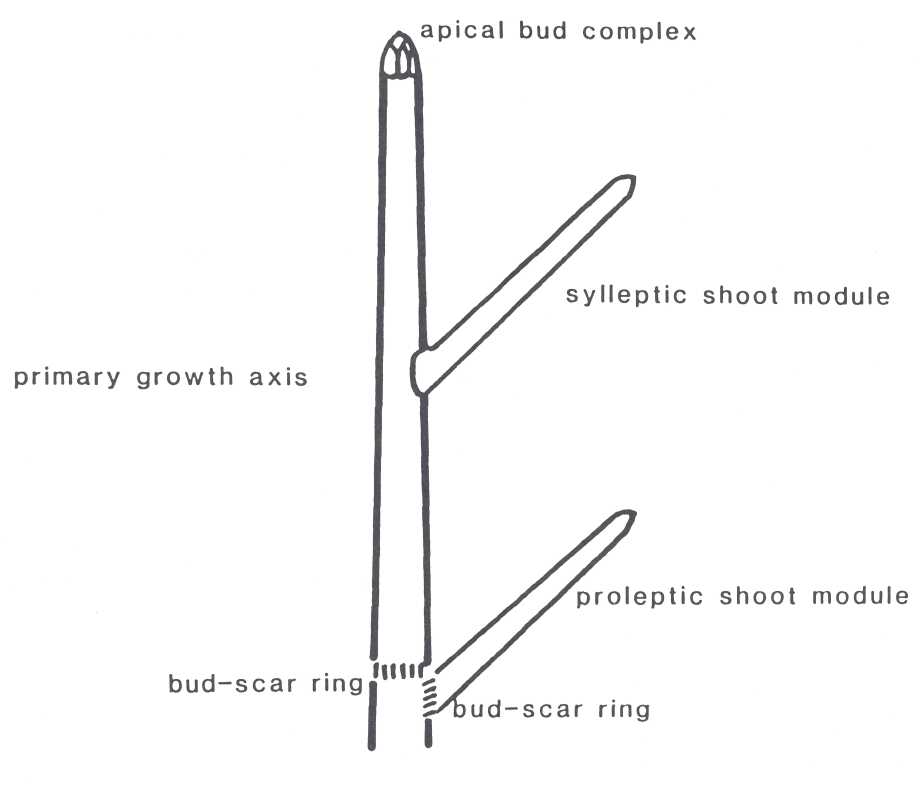
Fig. 1. Rhythmic growth module