Proc. of Second World Avocado Congress 1992 pp. 173-178
Boron:
Uptake by Avocado Leaves and Influence on Fruit Production
P.J.
(Hannes) Robbertse and Lou A. Coetzer
Margaretha
Mes Institute, University of Pretoria, Pretoria, 0002, Republic of South Africa
Ferdinand
Bessinger
Soil and
Irrigation Research Institute, Private Bag X79, Pretoria, 0001, Republic of
South Africa
Abstract. Absorption of boron by 'Hass' avocado leaves and the
influence of boron on fruit set and fruit production were studied. We found
that the absorption of boron by mature leaves was poor and that practically no
boron was absorbed by the adaxial leaf surface. Although not convincingly, we
found that fruit set and fruit production were improved by spraying trees with
Solubor (2 g/L) at the early stages of flowering. The results are discussed in
relation to the position of the tree along a slope in the experimental grove
and the influence of nitrogen on boron levels in the leaves.
Although the primary biochemical role of boron in plants is not yet fully understood (Teasdale and Richards, 1990), the influence of boron on different aspects of plant growth and reproduction is documented. It was demonstrated by Vasil (1963) that boron has a major influence on pollen tube growth and Robbertse et a/. (1990) found that boron also influences the directionality of pollen tube growth. For optimal pollen tube growth in avocados, the boron concentration in the flowers needs to be about 100 mg/kg (Robbertse and Coetzer, 1988). From this it can be derived that a certain concentration of boron in the plant is required prior to flowering for proper fertilization and fruit set. The question is, how much? Since the boron content in plants is traditionally determined by leaf analyses, one of the objectives of this study was to determine the boron content of avocado leaves prior to flowering and to relate this to fruit production. We also wanted to know whether the boron concentration in the leaves could be supplemented by spraying with a boron containing solution.
In this paper we report on the results obtained in field experiments in which the boron uptake by avocado leaves was monitored, as well as the effect of boron leaf spray on fruit production. The effects of age and position on the mineral composition of mango and avocado leaves was discussed by Koo and Young (1972, 1977), but we attempted to sample the latest mature leaves on flowering and fruiting branches to relate boron concentration with fruit set.
Materials and Methods
Two experiments were conducted on mature trees of the cultivar Hass.
Experiment 1. Two trees in full flush, growing on the experimental farm of the University of Pretoria, were treated with a 1 g/L Solubor solution as follows:
Tree 1: On one of the branches all the leaves were submerged in the solution. On a second branch, all the leaves were moistened only on the adaxial surface and on a third branch, all the leaves were moistened only on the abaxial surface.
Tree 2: This tree was used as a control and no treatment was applied.
Leaf samples from the young flush as well as mature leaves from the previous flush, were taken for boron analysis (A) before treatment, (B) one day after treatment, and (C) five days after treatment. The leaves were thoroughly washed in distilled water before drying.
Experiment 2. Mature trees in a commercial grove at the Westfalia Estate were used. Forty randomly chosen trees were divided into four groups on which different treatments were applied. In the first group (treatment A), the trees were sprayed once to saturation with Solubor (2 g/L) at the stage of inflorescence bud break. In the second group (treatment B), trees were sprayed twice with the same solution, first at bud break and again at apotheosis of the first flowers on the panicles. In the third group (treatment C), the trees were sprayed once at apotheosis of the first flowers in the panicle. In the control (treatment D), the trees were not sprayed. Leaf samples for boron analysis were taken at intervals from the youngest mature leaves on each tree. Leaves were washed in distilled water before drying. Samples of whole inflorescences from each tree were also taken for boron analysis. Fruit set was determined by marking ten flowering branch ends around the circumference of each tree and counting the number of fruit on these ends five months after flowering. The total fruit production per tree was also recorded.
Results
The results of experiment 1 are given in Figure 1. In the control samples the boron concentration of the young flush leaves is higher than that of the mature leaves. In all the treatments the boron uptake by young leaves was higher compared to that of the mature leaves. Compared to the control, practically no boron was taken up by the adaxial surfaces of the leaves.
The boron analyses of the leaves from experiment 2 are presented in Figure 2. The leaves sampled on 09/11/89 were from the previous summer flush, about five months of age, and contained very low boron concentrations. The Solubor spray of 06/25/89 had very little influence on the boron content. The spring flush leaves sampled on 11/28/89 had higher boron concentrations which were significantly affected by the Solubor sprays of 09/11/89. As the leaves aged until 06/14/90, the boron concentration decreased again. There were no obvious differences in the boron concentrations of leaves from the different treatments sampled on 08/16/90. These were four-month-old summer flush leaves and compared with the control, the Solubor spray of 06/14/90 had no effect. Contrary to the 1989 results, the boron concentration of the spring flush leaves sampled on 12/01/90, was not affected by the August Solubor spray.
Results showing the relationship of the boron concentration of the inflorescences, fruit set and fruit production are given in Figure 3. Boron concentration of the inflorescences was only determined during the 1989/90 season and samples were taken on 08/11/89, after the Solubor spray of 06/25/89. At this stage only treatments A and B were applied and both had significantly higher boron concentrations compared to the unsprayed treatments C and D. The fruit set results of the 1989/90 season were discarded due to poor sampling. Although not statistically significant, fruit production, determined on 08/11/90, was the highest in treatment A, which relates favorably with the boron concentration of the inflorescences. Fruit set for the 1990/91 season was the highest in treatments B and C which received the later Solubor spray. The reason why the results of the two seasons differ, may be ascribed to the fact that flowering during the 1990/91 season started a few weeks later than during the 1989/90 season, which means that the first spray in the first season was comparable with the second spray of the second season.
The
grove used for the field experiment was planted on a north-facing slope.
When comparing the experimental variables of the individual trees, we noticed
that the boron concentration as well as the fruit production of the trees at
the lower end of the slope seemed to be lower than that of trees from higher
altitudes. To test the validity of this observation, arbitrary lines were drawn
along the contours on a map of the grove in order to subdivide the experimental
trees into four new groupings. Each group of ten trees thus formed, included
even numbers of each treatment mentioned in experiment 2. Calculations were
made of all five boron analyses conducted during the two seasons and the
averages for each new combination are given in Figure 4 in relation to the
averages of the 1989/91 fruit production figures of the same trees. From this
comparison it is quite clear that the position of the tree along the slope had
a major effect on its fruit production and to a lesser extent on the boron
concentration of its leaves.
Discussion
Only a limited amount of boron is absorbed by mature avocado leaves. It is, therefore, not surprising that no significant differences were found in the boron concentration of leaves from the different treatments sampled on 09/11/89 (Fig. 2). However, there was a significant increase (LSD = 4.8) in the boron concentration of the inflorescences sampled on the same date (Fig. 3), which means that, either the boron was transported directly to the inflorescences, or the boron was absorbed by the inflorescences. This difference was carried through to the fruit production stage in treatment A, but not in treatment B (Fig. 3). The reason for this could be related to the position of the trees along the slope (Fig. 4). Fruit production was, however, severely affected by pronounced alternative bearing that occurred in both seasons. A fairly good relationship was found between the boron concentration of the leaves sampled on 06/14/90 (Fig. 2) and fruit set determined on 08/16/90 (Fig. 3). Fruit production figures for the latter season are still outstanding as the fruit are still on the trees.
In Figs. 1 and 2, we indicated that there was a reduction in the boron concentration of the maturing and ageing leaves. Labanauskas et al. (1958) showed that avocado leaves with a high nitrogen level contained significantly less zinc, copper and boron than leaves with less nitrogen. A nitrogen-zinc relationship in avocado leaves was also found by Lahav and Kalmar (1990). In another paper Labanauskas et al. (1961) reported a reduction of boron in ageing avocado leaves that was related to the nitrogen concentration in the leaves. In the first year of their experiment, the nitrogen level in the soil was high and the boron concentration in the leaves remained at a constant low level « 30 ppm). In the third year of their experiment, there was a decrease in the nitrogen level and the boron concentration in young leaves increased to between 50 ppm and 60 ppm before decreasing to 30 ppm in fully matured leaves. In our experimental grove, the nitrogen level in the soil was about 2%. This high level could explain the low levels of boron in figure 1 and the variation over the two seasons if our results are seen in relation to those of Labanauskas and his co-workers. The variation in the leaf boron concentrations and fruit production of trees along the slope (Fig. 4) may also be related to the leaching of nitrogen down the slope.
In a recent publication. Teasdale and Richards (1990) reported on the interaction of calcium and magnesium with boron. They highlighted the fact that an artificial boron deficiency can be created in cultured pine cells by increasing the level of magnesium in the growth medium. The influence of magnesium and calcium on the results of our field experiments can, therefore, not be excluded and will have to be considered in future experiments of this nature.
Literature Cited
Koo, R.C.J. and T.W. Young. 1972. Effects of age and position on mineral composition of mango leaves. J. Amer. Soc. Hort. Sci. 97:792-794.
Koo, R.C.J. and T.W. Young. 1977. Effects of age, position, and fruiting status on mineral composition of 'Tonnage' avocado leaves. J. Amer. Soc. Hort. Sci. 102:311-313.
Labanauskas, C.K., T.W. Embleton, and W.W. Jones. 1958. Influence of soil applications of nitrogen, phosphate, potash, dolomite, and manure on the micronutrient content of avocado Leaves. Proc. Am. Soc. Hort. Sci. 71:285-291.
Labanauskas, C.K., T.W. Embleton, W.W. Jones, and M.J. Garber. 1961. Seasonal changes in concentrations of zinc, copper, boron, manganese, and iron in 'Fuerte' avocado leaves. Proc. Amer. Soc. Hort. Sci. 77:173-179.
Lahav, E., Y. Bar and D. Kalmar. 1990. Effect of nitrogenous fertilization on the annual variations in nutrients in avocado leaves. Commun. In Soil Sci. Plant Anal. 21:1353-1365.
Robbertse, P.J. and L.A. Coetzer. 1988. The influence of boron on pollen germination, pollen tube growth and fruit set in some avocado cultivars (in Afrikaans). S.A. Avocado Growers' Assn. Yrbk. 11:65-67.
Robbertse, P.J. JJ. Locke, E. Stoffberg, and L.A. Coetzer. 1990. Effect of boron on directionality of pollen tube growth in Petunia and Agapanthus. S. A. J. Bot. 56:487-492.
Teasdale, R.D. and O.K. Richards. 1990. Boron deficiency in cultured pine cells: Quantitative studies of the interaction with Ca and Mg. Plant Physiol. 93:1071-1077.
Vasil, I.K. 1963. Effect of
boron on pollen tube germination and pollen tube growth. pp. 107-119. In:
Pollen physiology and fertilization. H.F. Linskens (ed.). North Holland,
Amerstand
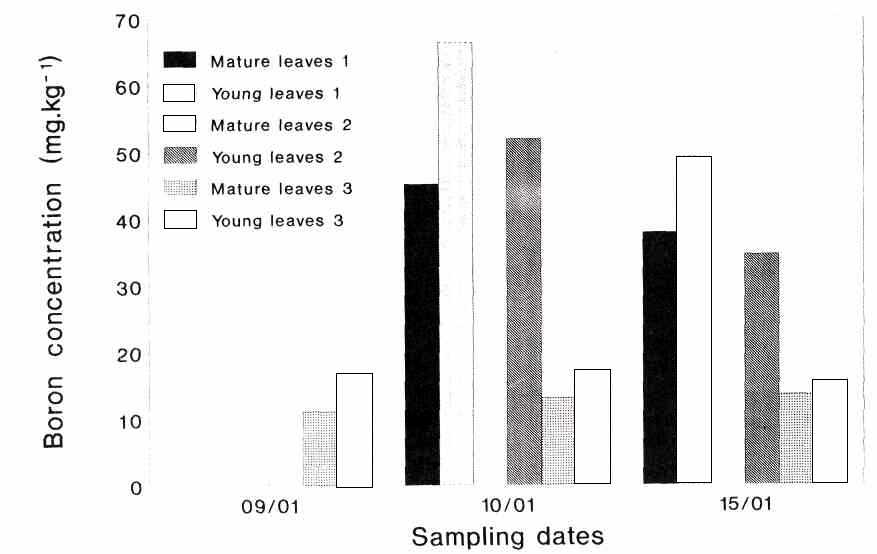
Fig.
1. Boron concentration in young flush and mature 'Hass' avocado leaves, one and
six days after treatments with Solubor (1 g/L with surfactant). Treatment 1:
Leaves submerged in Solubor. Treatment 2: Leaves moistened on abaxial surface. Treatment 3: Leaves moistened on adaxial
surface.
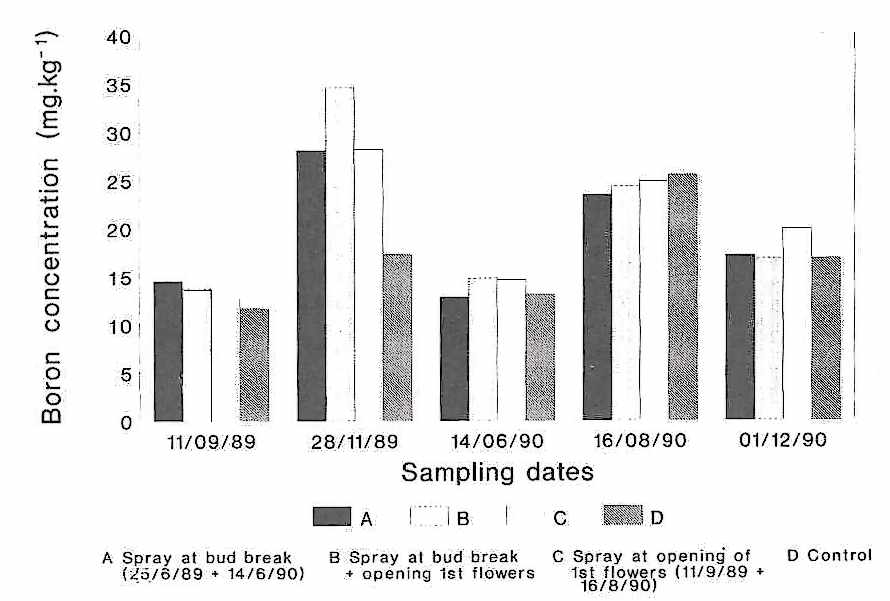
Fig. 2. Boron concentration of mature 'Hass' avocado
leaves from latest flushsampled at intervals after spraying with Solubor (2
g/L): A - at inflorescence budbreak (26/6/89 and 14/6/90); B - at bud break as
for A and again at anthesis of first flowers as for C; C - at anthesis 11/9/89
and 16/8/90).
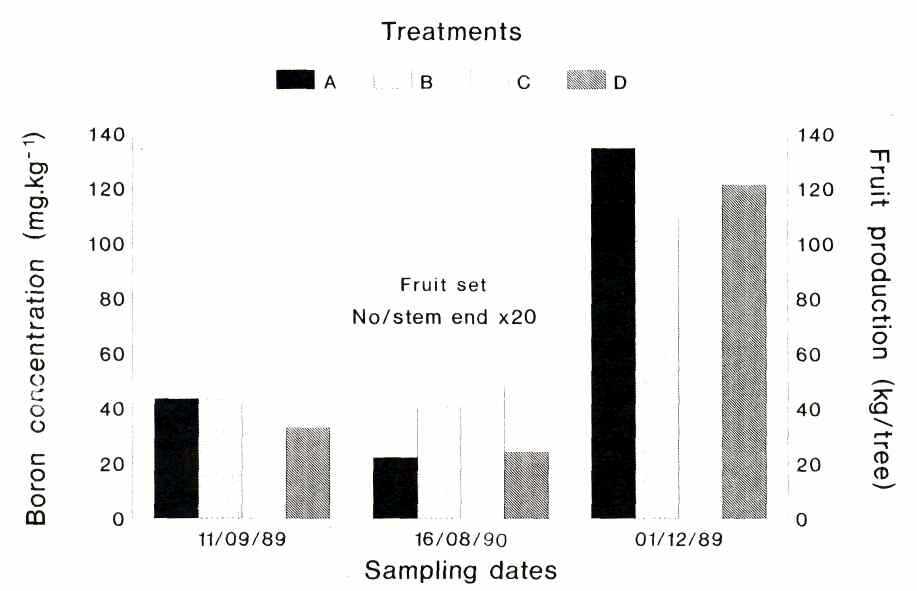
Fig. 3. Boron concentration of inflorescences, fruit set and fruit production of 'Hass' avocado sampled at different intervals after spraying with solubor (2 g/L): A - at inflorescence bud break (26/6/89 and 14/6/90); B - at bud break as for A and again at anthesis of first flowers as for C; C - at anthesis of first flowers (11/9/89 and 1 6/8/90); D - was not sprayed.
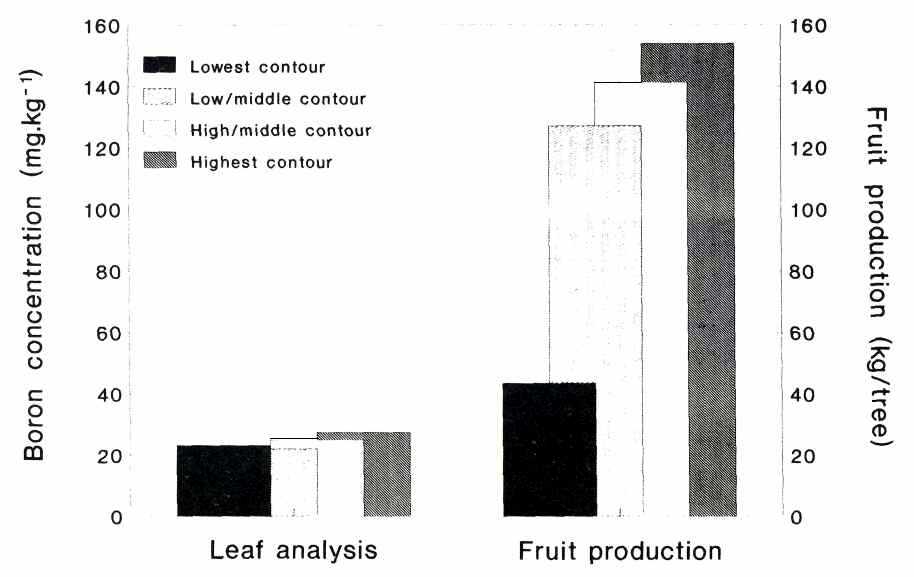
Fig. 4. Average boron concentration of mature 'Hass' avocado leaves sampled at intervals from latest flush over a period of two seasons and average fruit production per tree along contours in the grove.
.