Proc. of
The Performance of Rootstocks Tolerant to Root Rot Caused by
Phytophthora cinnamomi
under Field Conditions in
J.A. Menge, F.B. Guillemet,
Department
of Plant Pathology,
Abstract. A survey
was made of 16 avocado groves planted with rootstocks which are tolerant to
root rot caused by Phytophthora cinnamomi. All groves sampled were infested with P. cinnamomi. Eleven rootstocks were evaluated for
performance. All Phytophthora-tolerant rootstocks
performed better than Borchard, a Phytophthora-susceptible rootstock. Thomas
gave the best overall performance, followed closely by D9 and Barr Duke. Duke 7
exhibited moderate tolerance to Phytophthora
but provided a standard with which to rate other tolerant rootstocks. G755
did not perform well and many trees exhibited leaf chlorosis,
thin canopies and poor fruit set. Duke 7 performance could be correlated with
soil Mn,
and G755 performance could be correlated with soil Fe. All rootstocks exhibited
low amounts of P, Cu and Zn in their foliage, indicating that certain nutrient
sprays may assist in the establishment of Phytophthora-tolerant rootstocks.
Phytophthora root rot
of avocado (Persea
The best long-term solution for
controlling avocado root rot is the use of Phytophthora-tolerant rootstocks. Zentmyer began the search for Phytophthora-resistant rootstocks in the 1940's
and 1950's and this led to the selection of Duke 6 and Duke 7 varieties (Zentmyer, 1963). In 1975, Duke 7 became the first Phytophthora-tolerant rootstock to become commercially
successful. Since then extensive screening, selection and breeding by Zentmyer, Coffey, and Gabor has
led to a number of new and promising varieties (Zentmyer,
1978; Coffey, 1987b, Gabor, 1990). Many of these
varieties have now been in the field for 10 to 20 years. The purpose of
this study was to survey existing groves of Phytophthora-tolerant rootstocks growing in
Phytophthora-infested soil and to note their long-term performance.
Furthermore, an attempt was made to correlate Phytophthora-tolerant
rootstock performance with various soil factors.
Materials and Methods
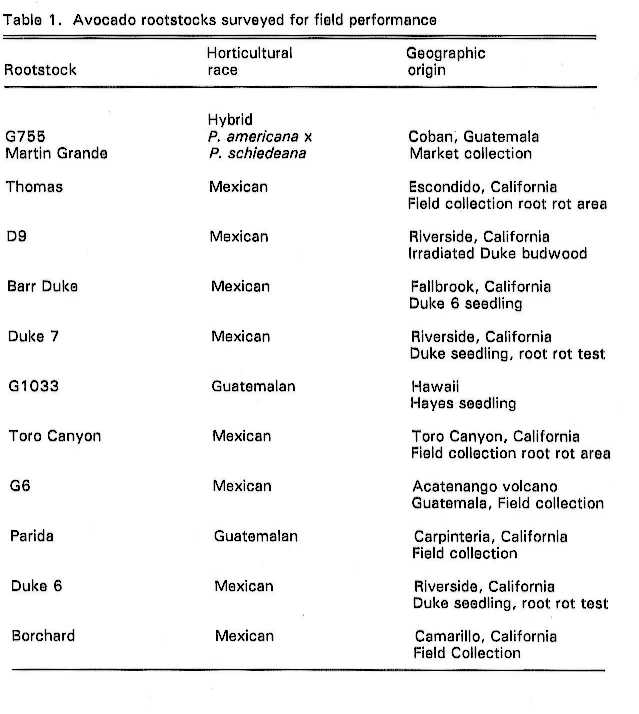
Sixteen avocado groves planted to Phytophthora-tolerant, clonal rootstocks were
surveyed January to June, 1990. All groves sampled had cv. Hass scions. All groves sampled had more
than one tolerant rootstock, which facilitated comparisons. The groves were
chosen to represent a cross section of the avocado industry in southern
Individual trees were rated by: 1)
visual rating of tree foliage on a scale of 0-5 (Gabor,
1990), where 0 = healthy and 5 = completely defoliated; 2) tree diameter
growth/year two cm above the bud union was calculated by measuring the diameter
and dividing by the tree age; 3) canopy volume growth/year was calculated by
measuring the height and diameter of the canopy using the formula for the
volume of an ellipse and dividing the result by tree age.
Soil samples were collected from the top 30 cm in the soil profile from ten trees in each grove. The soil was bulked and mixed for each grove. Soil samples for each grove were analyzed for total nitrogen using a semi micro-kjeldahl method (Black et al., 1965). The soil was digested in 97% KSO4 and 3% CuSO4.5H2O. Extractable P was determined using the bicarbonate (Olsen) method and quantified calorimetrically using the phosphomolybdenate complex (Chapman and Pratt, 1961). Exchangeable soil CA, Mg, K and Na were determined using atomic absorption spectroscopy following extraction with lithium chloride and lithium acetate (Yaalon et al., 1962). Soil Zn, Mn, Cu, and Fe were extracted using DTPA and quantified by atomic absorption spectrophotometry (Linsey and Norrell, 1969). Chlorine was detected in the saturation extract using a chlorodometer (Black et al., 1965). Percent soil organic matter was determined by the ignition method (Ball, 1964). Electrical conductivity (salinity) and pH were determined for each soil from a water saturation paste (Chapman and Pratt, 1961). Saturation % was the amount of water added to 100 g of soil which was necessary for saturation. Sodium absorption ration (CAR) was calculated with the formula
![]()
Soil composition Ca + Mg (clay,
silt and sand) was determined (Black et al., 1965).
Three to four leaves from 10 trees
per grove were gathered from groves containing G6, Duke 7, G755, and
Performance data was com- pared
using ANOVA and LSD analysis for data with unequal replications (Borland, Inc.,
1985). Soil and foliage nutrient characteristics were correlated with performance
values using regression (Impact Program).
Results
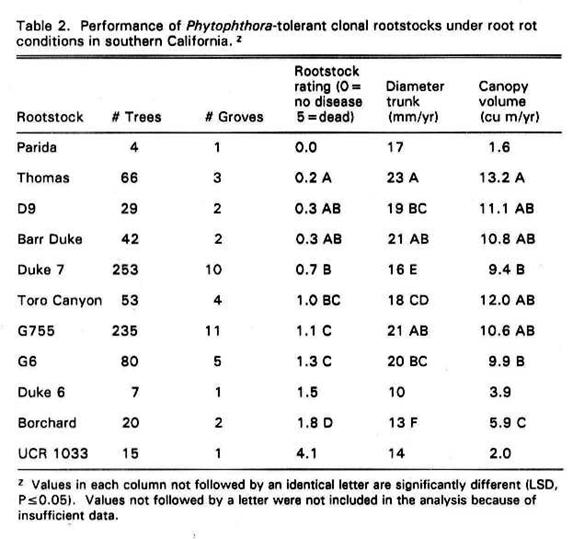
Although mortality was high in
some groves, in general the Phytophthora-tolerant rootstocks appeared
capable of surviving, growing and producing adequate fruit under optimal
growing conditions and optimal care. All of the resistant rootstocks performed
significantly better then Borchard, which is a Phytophthora-susceptible rootstock (Table 2). The rootstock which performed
the best under field conditions was Thomas. It was the leader in all
categories: foliage rating, trunk diameter/year and canopy volume/year (Table
2). D9 and Barr Duke also performed well in the field and were not
significantly different from Thomas in most categories. However, D9 appears to
establish slowly and young trees are often much smaller than other rootstocks.
This is shown by the fact that D9 had a significantly smaller trunk
diameter/year than Thomas.
G755 based on a visual rating did
not perform as well as Thomas, D9, Barr Duke, or Duke 7 (Table 2). G755 often
appeared to exhibit slightly chlorotic leaves and a
thin canopy. Poor fruit production was evident in many groves. However, growth
was vigorous and increases in trunk diameter/year and canopy volume/year were
not significantly different from Thomas. In general, G6 did not perform well in
the field, although values for trunk diameter/year and canopy
volume/year were similar to those of Duke 7 (Table 2).
Performance listings were included
for Parida, Duke 6 and UCR 1033 although the number
of trees examined for these rootstocks was not sufficient to be included in the
analysis (Table 2).
In an attempt to correlate root-
stock performance with soil conditions, we could find no correlations between
soil pH, salinity, total N, P, K, Ca, Na, Mg, Cu, Zn, Mn,
CI, saturation %, organic matter %, clay %, silt % or sand % and performance of
Thomas, Toro Canyon, Duke 7, G6 or G755. However both trunk
diameter/year and canopy volume/year for Duke 7 were positively correlated (P<0.10)
with soil Mn. All rootstocks, together, were
positively correlated with soil Mn (P<0.05).
Canopy volume of G755 was positively correlated (P<0.10) with soil Fe.
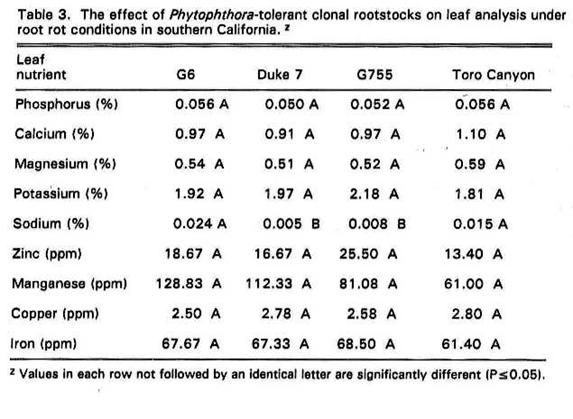
Performance of Thomas,
When leaf analysis was compared
for Thomas,
Discussion
In general results from this field
survey support and verify results from field and greenhouse work (Coffey,
1987a; Coffey and Guillemet, 1987; Gabor, 1990). All Phytophthora-tolerant varieties performed better
than the susceptible rootstock, Borchard. Duke 7
exhibited moderate tolerance to Phytophthora
root rot and may provide a standard with which to compare other Phytophthora-tolerant
rootstocks. Thomas gave the best field performance, closely followed
by D9 and Barr Duke.
The performance of D9 was somewhat
surprising. In previous field trials, the performance of D9 was somewhat variable
(Coffey, 1987a, Gabor, 1990), although Gabor found it to exhibit the highest level of resistance
to P. cinnamomi. It appears that D9 grows
relatively slowly when young, and it may not establish as well as more vigorous
rootstocks. However, the survey revealed that after 10 to 12 years, trees of D9
rootstocks were among the
largest trees in the survey.
G755 rootstock, although exhibiting
a very high level of tolerance to P. cinnamomi, did
not perform well in the field. The foliage often exhibited a slight chlorosis and the canopy was often thin. More disturbing
was the fact that fruit production was poor in many groves. The poor performance
of G755 is not thought to be due to lack of Phytophthora
tolerance, since poor performance evaluations were also made in fields
lacking Phytophthora. It is thought
that G755 requires additional iron fertilization, and this fact was verified by
the correlation in canopy volume/year and soil iron. However, in addition to
this problem, it is now believed that G755 suffers excessively from cold
temperatures (unpublished data). Since G755 parentage includes P. schiedeana, which is cold sensitive, it may be slightly
damaged each winter. This would affect visual ratings and perhaps fruit set
during the spring. Observations later in the summer and fall confirm that at
this time G755 appears greener and extremely vigorous.
Attempts
at correlating soil and foliage nutrient characteristics with performance of Phytophthora-tolerant avocados was not
highly successful. Much greater replications will be necessary to identify soil
factors which affect rootstock performance. Nevertheless, correlations between
soil Mn and Fe and rootstock performance indicate that
rootstocks should be evaluated carefully for effects on nutrient absorption as
well as Phytophthora resistance.
Foliar P, Zn, and Cu values for
all Phytophthora-tolerant varieties tested were quite low (Goodall et al., 1965). In addition soil Mn and Fe were found to affect performance in some rootstocks.
These elements do not diffuse readily in soil and Phytophthora
root rot may inhibit uptake of these elements. Supplemental fertilization
with these elements may enhance the performance of Phytophthora-tolerant rootstocks.
Literature Cited
Ball, D.F.
1964. Loss-on-ignition as an estimate of organic matter and
organic carbon in non-calcareous soils. J. Soil Sci.
15:84-92.
Chapman, H.D. and P.F. Pratt. 1961. Methods of analysis for
soils, plants and waters. Univ. of
Coffey, M.D. 1991. Cause and
diagnosis. Avocado root rot.
Coffey, M.D. 1987a. Phytophthora
root rot of avocado: an integrated approach to control
in
Coffey, M.D. 1987b. A look at
current avocado rootstocks.
Coffey, M.D. and F. Guillemet. 1987.
Profiles of UCR clonal rootstocks.
Gabor, B. 1990. Quantitative analysis of resistance to Phytophthora
cinnamomi in avocado rootstocks. Ph.D.
Thesis.
Goodall, G.E., T.W.
Embleton, and R.G. Platt. 1965.
Avocado fertilization. Leaflet.
Kotzé, J.M. and
J.M. Darvas. 1983. Integrated control of
avocado root rot.
Labanauskas, C.K.,
E.M. Nauer, and C.N. Roistacher. 1967.
Initial soil-mix and postplanting liquid
fertilization effects on nutrient concentrations in
Linsey, W.L. and
W.A. Norrell. 1969. Development of a DTPA
micronutrient soil test. Agron. Abst. 69:84.
Pegg, K.G.,
L.I. Forsberg, and A.W. Whiley. 1982. Avocado root rot. Qld. Agric. J. 108:162-168.
Yaalon, D.H.,
J.W. Schuylenborgh and S. Sluger. 1 962. The determination of cation-exchange characteristics of saline and calcareous
soils. Neth. J. Agric. Sci.
10:217-222.
Zentmyer, G.A. 1963. The Duke avocado.
Zentmyer, G.A. 1978. Origin of root rot
resistant rootstocks.
Zentmyer, G.A. 1980. Phytophthora
cinnamomi and the diseases it causes. Amer. Phytopath. Soc. Monograph 10, 96 pp.