|
- INTRODUCCIÓN
Debido a que el agua aportada en forma de riego localizado genera humedecimiento parcial de la superficie del suelo (BRESLER, 1977; TORO, 1995), el conocimiento de la distribución del sistema radical es importante para el manejo de riego y fertilizantes (VELLIDIS, SMAJSTRLA, ZAZUETA, 1990), ya que una alta densidad de raíces permite un mejor uso del agua y fuentes minerales (ATKINSON, 1980).
En el palto, la raíz puede ser considerada un sistema compuesto de raíces largas, raíces cortas y pelos radicales. Las raíces largas forman la mayor parte del sistema permanente, son generalmente leñosas y tienen crecimiento radial cambial. Las raíces cortas, a su vez, son de efímera existencia (máximo dos años), no tienen actividad cambial y al comienzo son tiernas y blanquecinas, pero luego se tornan suberizadas, de color pardo, conformando una parte importante del sistema radical (GIL, 1997).
Las raíces pueden desarrollar proyecciones de sus células epidérmicas denominadas pelos radicales que incrementan el contacto suelo-raíz, sin embargo, el palto presenta una baja frecuencia lo que lo hace ineficiente en la absorción de agua (WHILEY, 1990).
Las primeras técnicas de cuantificación de raíces se caracterizan por ser destructivas, laboriosas y alteran no sólo las raíces sino que además el sistema suelo que las contiene. Dentro de los antiguos métodos de investigación radicular se encuentran las excavaciones, el uso del barreno, perfiles de suelo y paredes de vidrio, desarrollados por WEAVER (1926); GHLAROV et al. , (1968); SCHUURMAN y GOEDEWAAGEN, (1971); KOLESNIKOV, (1971); BÓHM, (1979) y UPCHURCH y TAYLOR, (1990), citados por BOX (1996).
Hoy en día se han desarrollado modernas técnicas de investigación que minimizan las alteraciones del medio edáfico, pero que involucran un alto costo y dificultad de implementar a nivel de campo. Además, la mayoría de ellas son utilizadas para estimar la extensión del sistema radical de plantas en maceta y presentan restricciones para su uso en árboles frutales, al igual que no entregan información acerca de la distribución de éstas en el suelo. Las técnicas consideradas de "alta tecnología" permiten el estudio de las raíces "in situ" mediante paredes de vidrio y/o tubos de vidrio, y en ellas se puede incluir: rizotrones, capacitancia eléctrica, microdensímetros, resonancia magnética, minirizotrones, rizolaboratorios, cámaras de televisión de alta resolución y tomografías con rayos X (BOX, 1996).
En general, las técnicas de investigación de raíces buscan cuantificar el sistema radical basándose en un principio que determina directa o indirectamente la cantidad de éstas presentes en un área de suelo, en una planta o en un grupo de ellas.
Varios investigadores han encontrado diferencias en el desarrollo de raíces en función del volumen de suelo humedecido por el sistema de riego, lo que toma especial importancia bajo riego por goteo ya que el agua humedece un pequeño volumen de suelo y las raíces se adaptan rápidamente creciendo dentro de ésta zona sin presentarse efectos detrimentales para el árbol (FAUST, 1989).
Al respecto, VILLABLANCA (1994) señala que en un huerto regado por microaspersión el crecimiento de raíces está fuertemente determinado por el volumen y distribución del agua aplicada y UGARTE (1996) indica que en riego por goteo las raíces se concentran cerca de la zona de máxima saturación. Lo anterior concuerda con lo observado por SALAZAR y CORTES (1986) que trabajando con paltos adultos (Persea americana Mill. var Fuerte) encontró diferencias en la distribución horizontal de raíces bajo condiciones de suelo pesado y liviano, observando una distribución más uniforme en suelo liviano.
BIELORAI et al. (1981), en naranjos adultos bajo riego localizado, encontraron una correspondencia entre altas densidades radiculares y mayores volúmenes de agua de riego aplicados y BEVINGTON y CASTLE (1985); BIELORAI (1982); BIELORAI et al. (1981) y HILGEMAN (1977) señalan que en árboles de distintas edades una baja disponibilidad de agua en el suelo reduce el crecimiento radicular. Por otro lado, altas cantidades de agua provocan saturación disminuyendo el flujo de oxígeno hacia estratas inferiores (LAHAV y KALMAR, 1992), factor que predispone para el ataque de Phytophthora cinamomi (WHILEY et al. 1987; ZENTMYER, 1972). Al respecto, VALORAS et al. (1964), indica que las raíces de palto no crecen cuando la tasa de difusión de oxígeno es menor a 20 mg cm -2 min-1.
Considerando que en riego localizado se generan zonas de mojamiento que en el suelo concentran las raíces absorbentes del árbol, se podría suponer que existen puntos donde una mayor densidad de raíces absorbentes producirá un mayor consumo hídrico en un lapso determinado. Estos puntos corresponderían a "centros de consumo de agua".
Si lo anterior es válido, entonces se podría cuantificar indirectamente la cantidad y localización de las raíces de un árbol, detectando la ubicación y magnitud de los "centros de consumo de agua", mediante mediciones sistemáticas del contenido de humedad del suelo en diversas posiciones del volumen edáfico correspondiente. Entonces, el objetivo de éste trabajo es determinar si existe una relación entre densidad de raíces absorbentes y consumo de agua, bajo condiciones de suelo pesado y liviano, en dos tipos de riego, goteo y microaspersión.
2. MATERIAL Y MÉTODO
2.1. Ubicación y caracterización de la zona en estudio:
El estudio se realizó en el sector La Palma, Provincia de Quillota, V Región, Chile. El suelo pertenece a la serie La Palma, es de tipo sedimentario, profundo y origen coluvial; substratum de gravas y piedras con material intersticial de suelo, permeabilidad moderada, buen drenaje; plano, en posición de plano inclinado y microrelieve ligeramente ondulado (MARTÍNEZ, 1981). El clima corresponde al tipo mediterráneo (NOVOA et al, 1989).
2.2. Material experimental:
2.2.1. Caracterización de los sectores en estudio
Para realizar el estudio, se escogieron dos sectores en la Estación Experimental "La Palma" y dos en el Fundo "La Palma", diferenciados por el sistema de riego y tipo de suelo (Cuadro 1).
2.2.2. Caracterización de los árboles.
En cada sector se seleccionaron tres árboles adultos cv. Hass, injertados sobre franco; homogéneos en cuanto a tamaño, vigor, sanidad y carga frutal. La distancia de plantación de los árboles es de 10 * 10 m, con uno en quincunce (correspondiente a otra variedad), en la Estación Experimental, y de 9*9 en el Fundo "La Palma".
|
SECTOR |
TIPO DE RIEGO |
TIPO DE SUELO |
|
EXPERIMENTAL
"A"
ESTACIÓN FUNDO "A" FUNDO "B" |
Microaspersión, 2 emisores por árbol ubicados
sobre la Microaspersión, 2 emisores Goteo, 12 emisores/planta, Goteo, 12 emisores/planta, |
Suelo "pesado", con dos Suelo
"liviano", cuya Suelo "pesado", con dos |
2.2.3. Instrumentos de medición
2.2.3.1 Sonda de capacitancia
La
sonda de capacitancia, es un instrumento usado en la agricultura para medir
contenidos de agua en el suelo. Se basa en el principio de que distintos
contenidos de humedad responden a cambios en la constante dieléctrica de un
material. La mayoría de los elementos presentes en el suelo tienen una
constante dieléctrica entre 2 y 4. La constante dieléctrica del agua es 78. Por
lo tanto, al aumentar el contenido de humedad en el suelo, la constante
dieléctrica de la mezcla suelo-agua también se incrementa (TROXLER ELECTRONIC
LABORATORIES, 1995).
El instrumento consta de una unidad de control, la sonda y la guía de acceso (Anexo 1). La unidad de control es el componente electrónico que guarda las medidas y pruebas de calibración para luego transmitirlas a una impresora o computador. La sonda es el sensor que mide el contenido de humedad del suelo mediante electrodos. La guía de acceso es el componente que permite la entrada de la sonda con la precisión requerida a la profundidad específica (TROXLER ELECTRONIC LABORATORIES, 1995).
2.2.3.2. Sonda de neutrones
La sonda de neutrones modelo CPN 503DR HYDROPROBE mide el contenido de humedad del suelo u otros materiales. Para determinar el contenido de humedad en el suelo, la cápsula radioactiva de Americium-241:Beryllium, emite neutrones radiactivos en la zona de suelo a medir. La alta energía de los neutrones es moderada por la colisión con los átomos de hidrógeno presentes en el agua del suelo y al perder energía se convierten en neutrones termal izados que son detectados por el sensor de Helium -3. Mientras mayor sea la cantidad de neutrones termalizados detectados por la sonda mayor será el contenido de humedad presente en el suelo (BOART LONGYEAR COMPANY, 1995).
Al igual que la sonda de capacitancia, consta de una unidad de control, la sonda y la guía de acceso. El volumen de suelo representado en cada medición es aproximadamente esférico con un radio de 15 cm. (Anexo 2)
2.3. Metodología de trabajo:
2.3.1. Movimiento de agua en el suelo
Para determinar el delta de humedad generado por el movimiento de agua en el suelo después de un riego, se tomaron lecturas de humedad volumétrica entre riegos consecutivos en zonas sin raíces bajo riego por goteo y microaspersión, tanto en suelo pesado como liviano (Anexo 6).
2.3.2. Consumo efectivo de agua en el suelo
Para determinar el consumo efectivo de agua por las raíces (CE) se consideró el delta de humedad entre riegos consecutivos, medido en zonas con raíces, menos el movimiento de agua en el suelo, medido en zonas sin raíces (Anexo 7). El cálculo se realizó de acuerdo a la siguiente fórmula:
CE=(ef-ei)R-(ef-ei)sr
Donde:
ei = Humedad volumétrica inicial
ef = Humedad volumétrica final
R= Con raíces
sr = Sin raíces
Las mediciones se realizaron 24 horas después de un riego y 1 hora antes del riego siguiente, a 50 cm. de distancia desde el tronco, separadas 50 cm entre sí y en tres profundidades: a 20 y 40 cm en el sector del Fundo (A y B) con la Sonda de Capacitancia y a 20, 40 y 60 cm en el sector de la Estación Experimental (A y B) con la Sonda de Neutrones.
Las diferencias de profundidades medidas se debe a la naturaleza de los suelos, que en caso del Fundo presenta gran cantidad de piedras bajo los 40 cm limitando la entrada del tubo de acceso a mayores profundidades.
2.3.3. Instalación de tubos de acceso.
Los tubos se instalaron evitando compactaciones o bolsones de aire (TROXLER ELRCTRONIC LABORATORÍES, 1995) (Anexo 3). Por cada árbol se ubicaron 5 tubos de acceso, considerando una separación de 50 cm, paralelos a la línea de riego, a 50 cm del tronco y 50 cm de la línea de riego (Anexo 4). La presencia de raíces en la zona próxima al emisor es baja (VILLABLANCA, 1994) y no existe efecto de la orientación en la densidad de raíces absorbentes (UGARTE, 1996)
2.3.4. Densidad radical
Para determinar la densidad radical se abrieron zanjas de 60 * 75 * 300 cm, paralelas a la línea de riego (Anexo 5). Mediante una cuadrícula transparente de 480 cm2, se contó frente a cada tubo el número de raíces activas caracterizadas por tenor de color blanco, no suberizadas y menores a 2 mm de diámetro (ADATO Y LEVINSON, 1991) en tres profundidades: 20, 40 y 60 cm. (Figura 1). Luego se calculó la densidad radical como:
D.R = No de raíces
activas / 480cm2
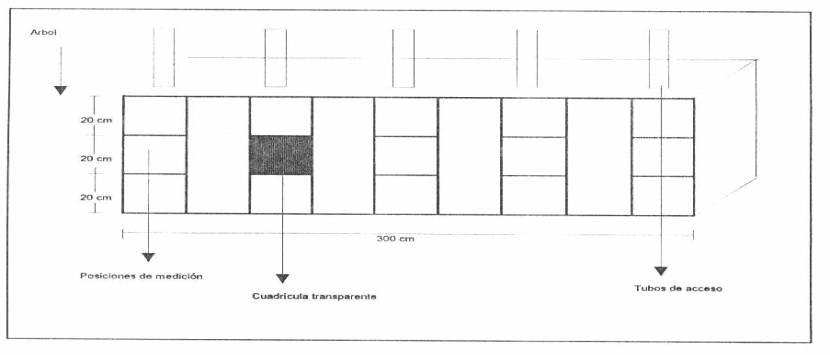
Figura 1. Posición de la cuadricula en la zona de medición.
3. PRESENTACIÓN Y DISCUSIÓN DE RESULTADOS
3.1. Análisis de densidad radical:
Al analizar la densidad de raíces presentes durante la temporada , se observó un notable descenso en comparación con lo observado por UGARTE (1996) y TORO (1995), tanto en el sector de la Estación Experimental "La Palma" como en el Fundo "La Palma" en mediciones realizadas en primavera y verano (Cuadro 2). En éste sentido, la falta de agua producto de un período de sequía que afectó a nivel nacional al sector silvoagropecuario, se tradujo en los árboles en estudio en una marchitez generalizada, clorosis, defoliación, exceso de floración y en una baja importante en la productividad por bajos calibres y caída de frutos. A nivel radical, es probable que gran parte de las raíces del árbol se trasladaran hacia estratas más profundas, encontrándose una baja cantidad de éstas en los primeros 60 cm de profundidad.
CUADRO 2. Densidad de raíces promedio (No raíces activas / cm2) por temporada y época de medición.
|
Temporada |
Primavera |
Verano |
|
94/95 |
1.64 |
3.72 |
|
95/96 |
|
|
|
97/98 |
0.009 |
0.007 |
3.1.1. Sistema de riego por goteo
De acuerdo a los resultados obtenidos en esta investigación, el consumo de agua generado por las raíces presenta una relación directa con la densidad de raíces a los 40 cm. de profundidad en suelo liviano y a los 20 cm en suelo pesado, con un coeficiente de determinación de 0,97 y 0,53, respectivamente.
Para las demás condiciones no se tuvo relación (Cuadro 3).
CUADRO 3. Análisis de regresión lineal para riego por goteo.
|
Suelo |
Profundidad |
Coeficiente |
Error |
F calculado |
F crítico |
|
Liviano |
0 a 20 |
0.08321989 |
0.00192548 |
0.02092154 |
0.89416357 |
|
Pesado |
0 a 20 |
0.53438264 |
0.00381765 |
1.19912123 |
0.35353796 |
|
Liviano |
20 a 40 |
0.97034157 |
0.00034187 |
48.3371308 |
0.006104 |
|
Pesado |
20 a 40 |
0.2340935 |
0.00462866 |
0.17393067 |
0.70468796 |
En general, el consumo hídrico en los primeros centímetros de
suelo no debería ser representativo de la densidad de raíces, ya que a este
nivel las pérdidas de agua por
evaporación directa son más significativas y afectan el abastecimiento de agua
por las raíces (KRAMER, 1989).
El sector regado por goteo correspondiente al Fundo "La
Palma", presentaba el momento de las mediciones una gran cantidad de hojas
en forma de "mulch" que
podría estar influenciando una menor pérdida de agua por evaporación.
VILLABLANCA (1994) encontró una mayor cantidad de raíces en suelos con “mulch”.
Lo anterior indica una influencia del "mulch" en el consumo hídrico
que debería considerarse al momento de determinar la densidad de raíces.
3.1.2. Sistema de riego por microaspersión
En microaspersión, los resultados se muestran inferiores comparados con los obtenidos en riego por goteo y sólo se observa una relación directa entre densidad de raíces y consumo hídrico en suelo pesado a los 60 cm. de profundidad, en cuyo caso el coeficiente de determinación es de 0,65 (Cuadro 4).
CUADRO 4. Análisis de regresión lineal para riego por microaspersión.
|
Suelo |
Profundidad |
Coeficiente |
Error |
F calculado |
F crítico |
|
Liviano |
O a 20 |
0.69235609 |
0.00126358 |
2.76210526 |
0.19510702 |
|
Pesado |
O a 20 |
0.340878 |
0.0025917 |
0.39442472 |
0.57453895 |
|
Liviano |
20 a 40 |
0.87655564 |
0.00155215 |
9.95056061 |
0.05108943 |
|
Pesado |
20 a 40 |
0.14145511 |
0.00356013 |
0.06125432 |
0.82049621 |
|
Liviano |
40 a 60 |
0.04003204 |
0.00294156 |
0.00481541 |
0.94904324 |
|
Pesado |
40 a 60 |
0.65186283 |
0.00236588 |
2.21671219 |
0.23327427 |
Para la condición de suelo liviano entre los O y 40 cm. de
profundidad se presenta una relación inversa entre consumo hídrico y densidad
radical bajo riego por
microaspersión. En este caso es probable que las mediciones del contenido de
humedad del suelo realizadas dos semanas antes del recuento de raíces no sean
representativas del consumo de agua generado por las raíces debido a la corta
vida que éstas presentan. Al respecto, FLORES, VITE y BORYS (1988) y UGARTE
(1996) señalan que las raíces finas del palto presentan un desplazamiento
ondulante de formación, regeneración y muerte que puede ser consecuencia de un
mecanismo
adaptativo del palto al constante cambio de las condiciones del suelo.
Por otro lado, bajo microaspersión se utilizan bajas frecuencias y
láminas de riego mayores en comparación a riego por goteo, lo que crea
problemas de aireación los
días siguientes al riego. Se encontró bajo riego por goteo un 44% más de raíces
absorbentes, similar a lo observado por UGARTE (1996) y TORO (1995) en el
mismo suelo, donde obtuvieron un 43% y
39%, respectivamente. Bajo estas condiciones, es posible que la muerte parcial
del sistema radical o el retraso en la
emisión de raíces blancas por condiciones de saturación del suelo, dificulte la
determinación de la densidad de raíces del palto por medio del consumo hídrico
cuando ambos parámetros no se miden simultáneamente. Bajo microaspersión en
suelo liviano, la alta conductividad hidráulica genera descensos en el
contenido hídrico por movimiento de agua en profundidad, tal como se muestra en
mediciones de humedad volumétrica realizadas entre riegos consecutivos en zonas
sin raíces (Figura 2). En suelo pesado, en cambio, se presentan condiciones de
sobresaturación que limitan el desarrollo de raíces absorbentes o aumentan la
posibilidad de problemas fungosos (LAHAV y KALMAR , 1983; WHILEY et al, 1987;
VILLABLANCA, 1994 y TORO, 1995). Lo anterior indica que los primeros 20 cm de
suelo pueden no ser representativos del consumo de agua generado por las raíces
absorbentes del palto. Al evaluar el contenido de humedad entre riegos consecutivos
en zonas sin raíces, se observó una influencia del sistema de riego y el tipo
de suelo en el movimiento de agua en el suelo (Anexo 6). En este sentido, el
movimiento de agua en el suelo puede ser un punto crítico en la determinación
del consumo de agua por parte de las raíces absorbentes del palto. Con respecto
a este punto se debe considerar que dentro de cada clasificación de suelo,
pesado y liviano, puede existir gran variabilidad de las características
físicas de éste en cuanto a textura, estructura y proporción de macro y micro
poros. Las gran pedregosidad del sector
en estudio presentan dificultad para la determinación el consumo hídrico
efectivo, ya que disminuye la capacidad de retención de agua del suelo y el
consumo hídrico queda "enmascarado" en un gran delta de humedad
generado por percolación del agua hacia estratas más profundas. La figura 3,
muestra el contenido inicial y final de agua entre riegos consecutivos y la
densidad de raíces presentes en el suelo bajo riego por microaspersión, y se
observa que en zonas sin presencia de raíces se generan deltas de humedad del
orden del 3% de humedad volumétrica. Del mismo modo, sectores de suelo con baja
densidad radical pueden generar deltas de humedad mayores que sectores con alta
densidad radical cuando la proporción de macroporos se incrementa por efecto de
la pedregosidad. Por otro lado, el sector bajo riego por goteo, presenta una mayor pedregosidad y los deltas
de humedad generados en sectores sin raíces superan el 10% de humedad volumétrica
(Figura 4).
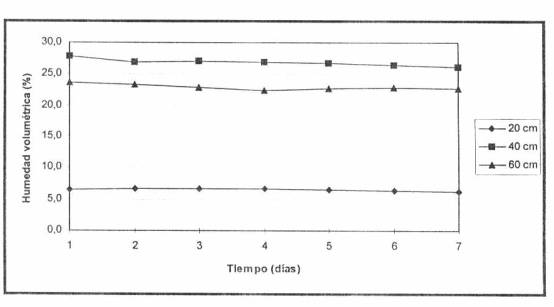
Figura 2. Humedad volumétrica diaria entre dos riegos bajo microaspersión, suelo liviano.
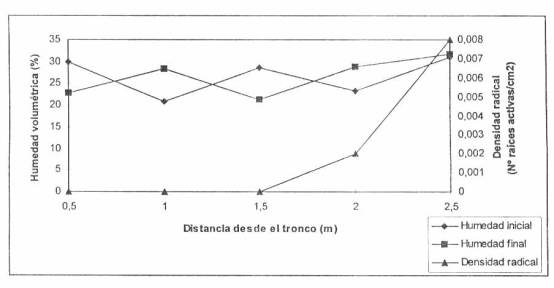
Figura 3. Densidad de las raíces absorbentes de palto bajo riego por goteo
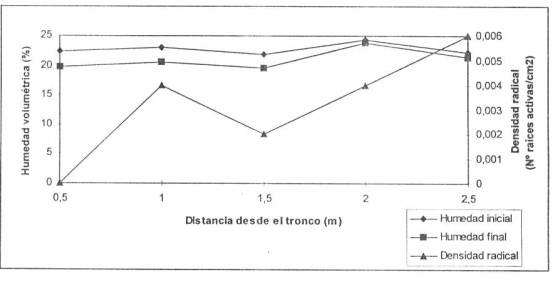
Figura 4. Densidad de raíces absorbentes de palto bajo riego por microasperción.
4. CONCLUSIONES
En el presente estudio se encontró una relación directa entre
densidad de raíces absorbentes y consumo hídrico bajo riego por goteo en
condiciones de suelo liviano a
140 cm de profundidad, suelo pesado a 20 cm de profundidad y riego por
microaspersión en condiciones de suelo pesado a 60 cm de profundidad.
La gran pedregosidad del sector en estudio dificulta la
determinación del consumo hídrico, ya que se generan diferencias de humedad
entre riegos consecutivos muy
grandes, "enmascarando" la pérdida de humedad del suelo por efecto
del consumo hídrico.
5. RESUMEN
El estudio de raíces
"in situ" ha sido desarrollado por varios investigadores, en
un comienzo con métodos laboriosos y destructivos para la planta y su medio
edáfico, y
actualmente con técnicas costosas y difíciles de implementar a nivel de campo
especialmente en árboles frutales. Una relación directa entre densidad de
raíces
absorbentes y consumo de agua podría servir para determinar el patrón de
distribución del sistema radical midiendo el consumo hídrico de las raíces.
Para determinar si existe relación entre densidad de raíces
absorbentes de Persea americana Mill cv. Hass y consumo hídrico se
realizó un estudio en la localidad de
Quillota bajo dos sistemas de riego, goteo y microaspersión, en condiciones de
suelo pesado y liviano.
La densidad de raíces absorbentes se obtuvo mediante el conteo de raíces activas en seis árboles adultos por sistema de riego, a tres profundidades de suelo, en una zanja paralela a la línea de riego.
El consumo hídrico se determinó midiendo la humedad volumétrica del suelo después de un riego y antes del riego siguiente. Como resultado de esta investigación no se encontró una relación directa entre densidad de raíces absorbentes y consumo hídrico, dado que la gran pedregosidad del sector afecta la determinación del consumo de agua en el suelo. No se descarta que bajo otras condiciones de suelo pueda comprobarse una relación directa entre los parámetros en estudio.
6. LITERATURA CITADA
ADATO, Y. and LEVINSON, B. 1991. Influence of Reduced Rates of Water and
Fertilizer Application Using Daily Intermittent Drip Irrigation on the Water
Requirements, Root Development and Responses of Avocado trees (cv. Fuerte).
Journal of Horticultural Science 66(4): 449:463.
ATKINSON, D. 1980. The distribution and
effectiveness of the roots of the crops. Hort. Rew. 2: 424-490.
BEVINGTON J.B. and CASTLE, W.S. 1985. Annual
root growth pattern of young citrus trees in relation to shoot growth, soil
temperature, and soil water content.
J. Amer. Soc. Hort Sci 110: 840-854.
BIELORAI, H; DASBERG, S; ERNER, Y. and BRUM, M.
1981. The effect of various soil moisture regimes and fertilizer level of
citrus yield under partial
wetting of the root zone. Proc. Int. Soc. Citriculture Cong (Tokyo) 1:585-589.
BIELORAI, H. 1982. The effect of partial
wetting of the root zone on yield and water use efficiency in a drip and
sprinkler irrigated mature grape fruit grove.
Irrig. Sci 3: 89-100.
BOART LONGYEAR COMPANY. 1995. Manual of operating 503 DR Hydro probe
Moisture Gauge. California, sp.
BRESLER, E. 1977. Trickle-drip Irrigation: principles and application to
soil-water management. Ad. In Agronomy 29:343-393.
BOX, J. 1996. Modem methods for root
investigations. In: Y, Waisel; A. Eshel; U. Kafkafi. Eds. Plant Roots. New
York, The Hidden Half. pp. 193-238
FAUST, M. 1989. Physiology of temperate zone
fruit trees. New York, John Wiley. 338 p.
FL FLORES, D; VITE, L and BORYS, M.W. 1988. Laterals distribution on the principal root in avocado seedlings (Persea americana Mill). California Avocado Soc. Yrb 72: 237-242
GIL, G. 1997. El potencial productivo. Crecimiento vegetativo y diseño de huertos y viñedos. Santiago, Ediciones Universidad Católica de Chile. 342 p.
H ILGEMAN, R.H. 1977. Response of citrus to water stress in Arizona. Proc. Int Soc. Citriculture Cong. (Orlando) 1: 70-74.
KRAMER, P.J. 1989. Relaciones hídricas de suelo y plantas. Una
síntesis moderna. México. Harla.
538 p.
LAHAV, E. and KALMAR, D. 1992. Water
requirements and the effect of salinity in an avocado orchard on clay soil.
Word Avocado Congress II Proceedings Orange, California, April 21 - 26. pp.
323-330.
MARTÍNEZ, A.R. 1981. Proyecto
de implementación de un sistema de riego tecnificado en la Estación
Experimental "La Palma", Quillota. Tesis Ing. Agr.
Quillota, Universidad Católica de Valparaíso, Facultad de Agronomía. 102 p.
NOVOA, R.; V1LLASECA, R.; DEL CANTO, P.; ROANET, J.; SIERRA, C. y DEL POZO, A. 1989. Mapa agroclimático de Chile. Santiago, INIA. 221 p.
SALAZAR, S. and CORTES, J. 1986. Root distribution of mature avocado tress growing in soils of different texture. California Avocado Society Yearbook 70:165-174.
TORO, M. 1995. Efecto del método de riego en la distribución
espacial del sistema radical de paltos (Persea
americana Mill.) cv. Hass, en dos tipos de suelos
Tesis Ing. Agr. Quillota, Universidad Católica de Valparaíso. 27 p.
TROXLER ELECTRONIC LABORATORIES, INC and
Subsidiary Troxler International LTD. 1995. Manual of operation and instruction
Sentry 200-AP. NC. EE.UU. sp.
UGARTE, M. 1996. Efecto del método de riego en
la distribución espacial del sistema radical de
paltos (Persea americana Mill.) cv.
Hass, en suelo franco
arcilloso. Tesis Ing. Agr. Quillota, Universidad Católica de Valparaíso. 33 p.
VALORAS, N; LETEY, J.; STOLZY, LH. and FROLICH,
E.F. 1964 The oxygen requirements for root growth of tree avocado varieties.
Proc. Amer. Soc. Hort.
Sci. 85: 172-178
VELLIDIS, G.; SMAJSTRLA, A. and ZAZUETA, F.
1990. Soil water redistribution and extraction patterns of drip irrigated
tomatoes above a shallow water table.
Transaction of ASAE 33 (5): 1525-1530.
VIllablanca, Y. 1994. Estudio comparativo de la distribución y densidad de
raíces absorbentes en palto (Persea
americana Mill.) cv. Hass en función a los
patrones de distribución uso-consumo del agua en el suelo evaluados bajo dos
sistemas de riego presurizado (goteo y microaspersión). Tesis Ing. Agr.
Quillota Universidad Católica de Valparaíso. 108p.
WHILEY, A.W.; PEGG, K.G.; SARANAH, J.P.B. and
LANGDON, P.W. 1987. Influence of
phytophthora root rot on mineral nutrient concentrations in avocado leaves.
Australian Journal of Experimental agriculture 27: 173-177.
ZENTMEYER, G.A. 1972. Avocado root rot. California Avocado Society
yearbook Vol. 66: 29-36.