ON AVOCADO FRUIT SIZE
D. N. Zamet
ACCO Experimental Station, Ministry of Agriculture,
The text of
this article was published in the 1995 Yearbook. Through misadventure, the
tables and figures intended for inclusion in the paper were omitted,
regrettably diminishing its usefulness. The paper is here presented in full,
inclusive of the graphic accompaniments provided by the author.
Summary
Data are presented to show that avocado fruit, both parthenocarpic and normal seeded fruit, show multimodular fruit size distribution. Attention is drawn to
the fact that with regard to the time of the flowering period of a cultivar the
peaks of fruit size and minimum temperature are very similar in number. It is
suggested that the minimum temperature controls the time and amount of fruit
set. It is also suggested that parthenocarpic fruit
are those fruitlets which though not having completed
the conditions for normal fruit development can, due to the in-tree hormone
status, still manage to "hang on" and grow.
Avocado fruit vary enormously in size (1) from less
than 10 grams to well over one kilogram (personal observation). This remarkable
size variation is usually between different cultivars; however, it also occurs
between fruit of the same cultivar, usually to a smaller extent; e.g., Fuerte, where the size can range from 20 grams to over 500.
Although some cultivars can give normal seeded fruit of very small size, most
of the smaller fruits are seedless and usually shaped like—and
called—"cucumbers" or "cukes" (in
a few cultivars these "cukes" are
spherical). Many people have labored very hard to explain the reason for these
seedless fruits.
On the way, they have produced excellent diagrams
describing the differences between seedless and normal fruit (3,12). Trees have been identified which give parthenocarpic fruit every year, and here differences in
floral behavior have been noted (11). Steyn et al. have suggested (11) "that
the disturbance of the polarity along the micropylar-chalazal
axis of the nucleus primordium sets in motion a
series of aberrant phenomena in the developing ovule." This prevents the
seed developing and can result in a "cuke."
This explanation, however, leaves open the question of "what causes the
disturbance?"
It is very difficult to know when a
particular flower sets and begins its way to becoming a fruit, normal or parthenocarpic. On a medium-size tree there will be
20—30,000 flowers open on average each day during the flowering period
according to counts made in 1976 (5), and of these a very small percentage
become fruit.
The occurrence of parthenocarpic
fruit varies from year to year, just as can the levels of normal fruit. It is
the author's opinion that for any particular cultivar, as the number of normal
fruit rises, that of the seedless goes down. Years of relatively low yields
seem to be the best years for cuke development. Steyn et al. (11)
found that if normal fruit did occur on "parthenocarpic"
trees", then they did not occur on the same branch. This author has found
them growing in the same inflorescence cluster on parthenocarpic
trees and many cases of Siamese twins—one normal and one seedless fruit growing
on the same fruit stalk (a two-ovuled flower). This
would tend to belay the idea that there is always something special about
particular flowers, branches, or trees; and that this speciality,
if present, can give rise to parthenocarpic fruit.
Zilka and Klein have shown that larger Hass fruit set
before smaller fruit (14). Sedgely and Annells have shown in the laboratory the importance of
temperature on avocado fruit set (14). Lomas showed
that heat stress during flowering and fruit set in the orchard played a role in
yield levels6. This author has shown that minimum temperature below 10°C
affects the crop level (13). Working together, Lomas
and Zamet have shown that crop level in avocado
depends on heat stress, minimum temperature, March soil temperature, and winter
rainfall (17). These four factors can be explained to work in the following
manner. Excess winter rainfall leads to death of many roots, so that in early
spring, during the flowering and fruit set period, the tree has difficulty in
absorbing sufficient moisture. Low soil temperature in March prevents root
growth immediately previous to this same critical period with similar effect of
insufficient moisture absorption. Low minimum air temperature prevents or slows
down the fruit set process, or causes the "disturbance" as suggested
by Steyn et al. (11) Heat stress removes all those fruitlets
which for one reason or another are "weak", or in extreme cases will
simply burn off the fruitlet. Total winter rainfall
and soil temperature are factors which do not change significantly from day to
day. Temperature, however, can fluctuate considerably in
Methods
As part of an investigation into fruit size,
fruit was picked from trees of several different cultivars, each bearing
numerous parthenocarpic fruit. Some of the cultivars
also bore normal seeded fruit which was also picked. Each fruit was weighed
separately. Normal fruit were weighed on a spring balance to the nearest gram,
while the parthenocarpic fruit were weighed on a Precisa electronic scale to the nearest tenth of a gram.
The weights were grouped into 20 gram groups for normal fruit and into two gram
groups for the parthenocarpic fruit. The percentage
of fruit in each group was calculated and distribution curves drawn.
The fruit examined was from the 1994-95 crop from trees growing on the Acco
station. In most cases, the total crop of the tree was picked within ten days,
the two exceptions being two trees of the cultivar 4403, where 271 fruit were
picked from a tree bearing an estimated 1,600 parthenocarpic
fruit, and 211 were picked from a tree bearing an estimated 600 parthenocarpic fruit. Both trees had two normal fruit each.
The minimum temperature regime data were
obtained from the records of the meteorological station close to the orchard.
These data were drawn as a graph for each day from the end of March through
till mid-May. In order to give an idea of how the temperature regime varies
between years in the flowering period, data were extracted from the records and
the information drawn as three day averages. Flowering data were also extracted
from the records.
Results
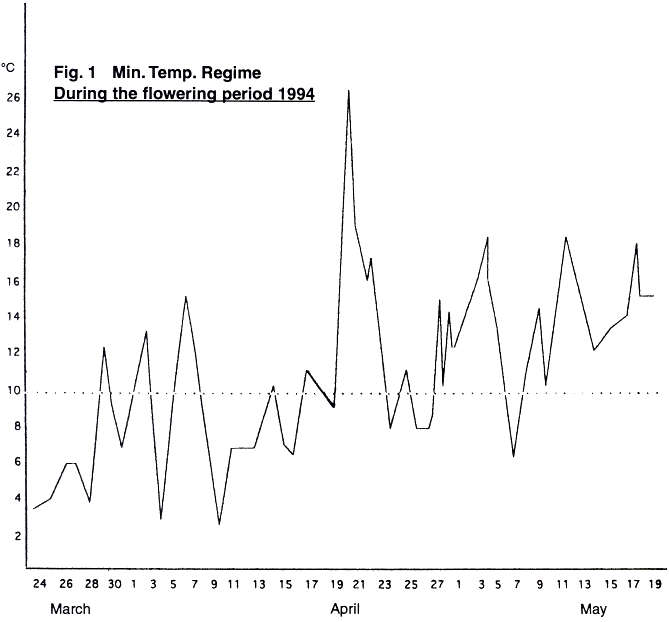
The minimum temperature regime for the
flowering period of 1994 is shown in Fig. 1. The fact that the minimum
temperature varied considerably from day to day can be clearly seen. It can
also be seen clearly that the minimum temperature varies frequently above and
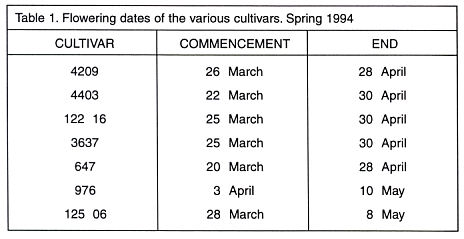
below the 10°C level, with maximum range of over 20 degrees. Fig. 2 shows the variation
in temperature regimes for selected years. The fact that the regimes vary from
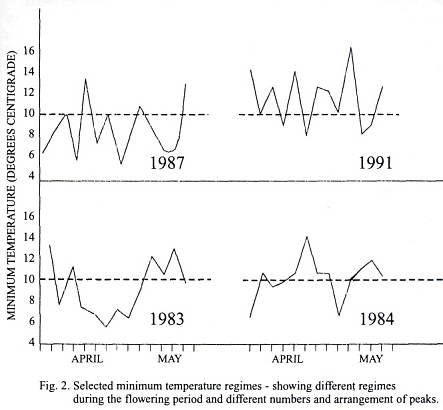
year to year can be seen, and also the overall level of the temperature. The
flowering period for each cultivar is shown in Table 1. These dates are
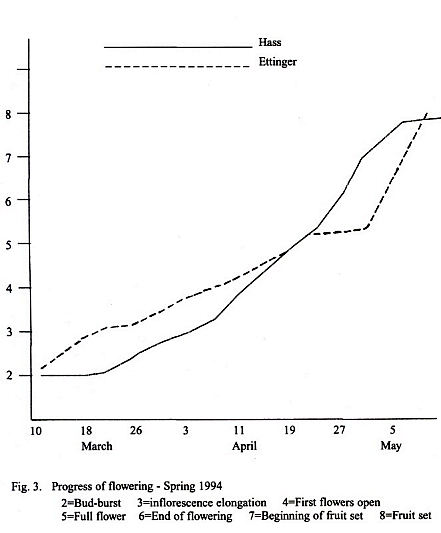
accurate to ±3 days. In Fig. 3 is given the progress of flowering for the
commercial cultivars Hass and Ettinger, showing the
relatively regular progress of flowering for 1994.
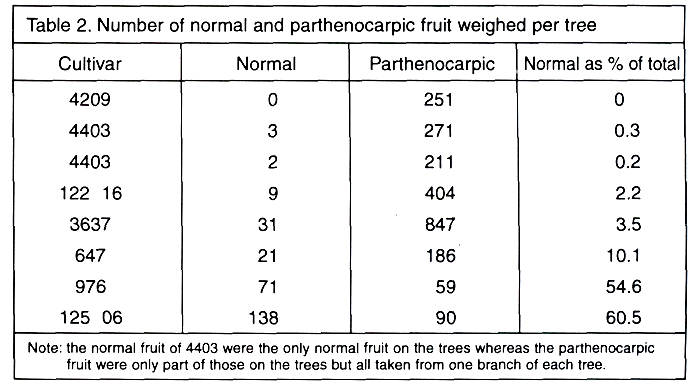
The numbers of fruits weighed from each tree
are given in Table 2. A total of over 2,000 fruits were weighed for this part
of the investigation.
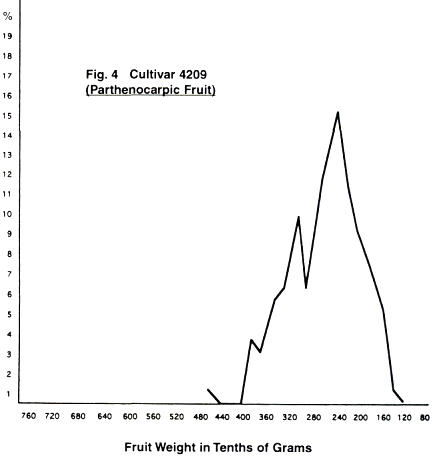
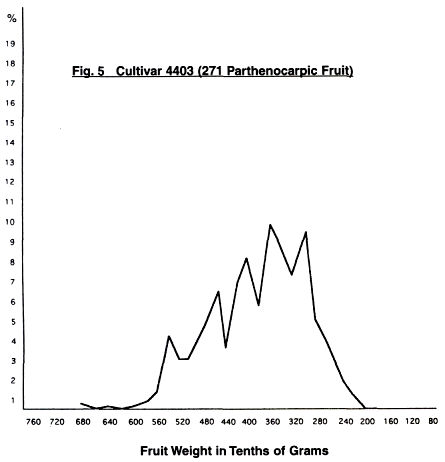
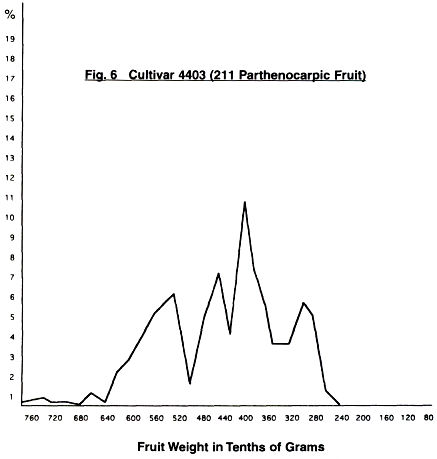
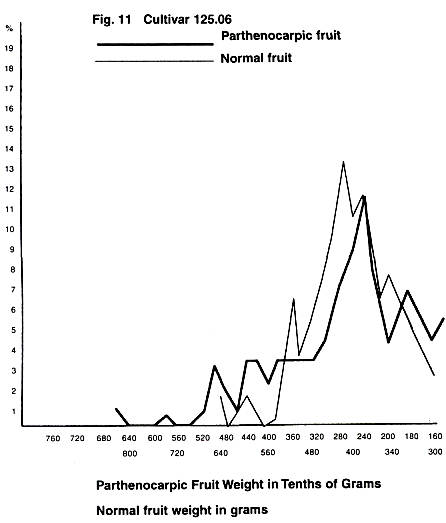
In Figs. 4-11 are given the fruit weight
distribution curves for the various cultivars. In three cases, distribution
curves for normal fruit are also given.
Discussion
Distribution curves are usually described as
"unimodal" or "multimodal"2. When
a multimodal curve is obtained, this indicates that more than one factor is
controlling the curve. Flowering is basically a unimodal
curve. It commences slowly, builds up to a maximum, and then falls off till
complete cessation. It can show slowing downs or speeding ups with prolonged
temperature variations7 and, as can be seen in Fig. 2 with the Ettinger cultivar; thus (except for a cultivar such as
Pinkerton, which can begin flowering in the autumn and give bursts of activity
as winter weather conditions permit), avocado flowering is basically unimodal.
If the work of Zilka
and Klein (14) is true, then fruit size should show a unimodal
distribution curve. The first set fruit will be the largest; however, as
flowering is in the beginning weak, the numbers of fruit setting will be few.
As flowering intensity increases, more fruit will be able to set, and this will
raise the curve; and then as flowering subsides, fewer fruit will be able to
set and the size distribution curve will fall.
The suggestion is often raised that citrus
and/or wild flowers act as competitors for the services of the honeybee. As far
as citrus is concerned, this frequently stops flowering quite suddenly
following a heat wave. If this would affect the avocado fruit set, then it
should be able to find a sudden single change in the distribution curve. In all
the work carried out so far, both on parthenocarpic
and normal fruit, this does not seem to be so. The trees used for this
investigation grow quite close to citrus, with the bee hives closer to the
citrus.
Temperature affects bee activity (4) and
also the opening and closing of the various stages of the flower (4). This
article does not purport to explain by what means minimum temperature affects
fruit set, and therefore the fruit size distribution curve, but it tries to
show that this is an actual possibility.
The minimum temperature regime during the 1994
flowering period (Fig.1) shows a typical situation for a
When we examine the fruit size distribution curves of
the cultivars used in this work, they seem to fall into several different
types. First, there are those which have a considerable crop of normal fruit in
addition to the parthenocarpic ones. It is interesting
to note that the two cultivars with considerable normal fruit carried on
flowering into the month of May, while the others finished by the end of April.
It has already been noted by other workers that late flowering cultivars are
frequently good yielders; and, on this basis, work is
being carried out to try to delay flowering in order to increase yield (9).
Another grouping can be made with cultivar 3637 on one side as compared to all
the others. The majority show multimodal distribution both
for parthenocarpic and for normal fruit, while 3637
shows multimodal distribution only with the normal fruit, and these are
very few. It would seem that under
When we compare the curves of the two 4403 trees, we
can see—although of different intensity—the same five peaks. Four occur as the
curve rises to a maximum, and then one which is smaller. There is a slight
indication of a sixth peak at the large fruit end, but this is less than one
percent of the total.
With the cultivar 4209, we have only four clear
peaks, but with a "bulge" around the 20 gram weight, and another
smaller one at the 34 gram weight, both of which could be signs of two weak
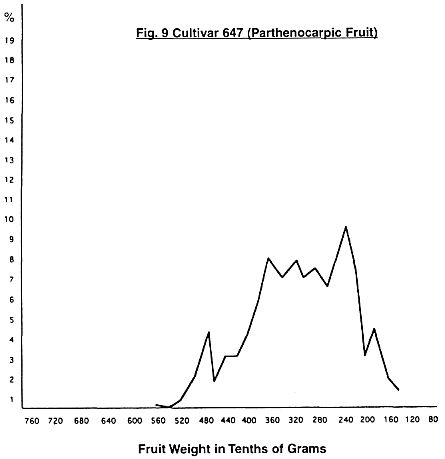
peaks giving a total of six peaks. In the case of cultivar 647, we have a wide
spread of weights and six clear peaks with a possible seventh at 44 grams. As
with 4403, we have one peak only on the downward run to smaller fruit. In the
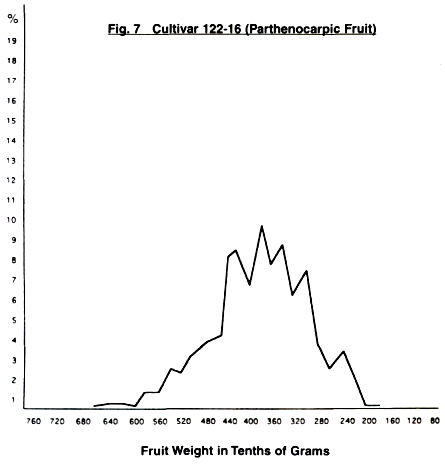
case of 122.16, we again have a wide distribution curve and five peaks as in
4403, but three of them are on the downward side as the fruit size drops.
With the two cultivars with a considerable crop of
normal fruit, we have eight or nine peaks with parthenocarpic
fruit and seven or nine with the normal fruit. In both cultivars, there is a
gradual increase in fruit size before reaching the maximum percentage and
several peaks on the downward run as the fruit gets smaller. There is a
considerable similarity between the distribution curves of the normal and of
the parthenocarpic fruit which again indicates the
strong possibility that the same factor is controlling the fruit size
distribution.
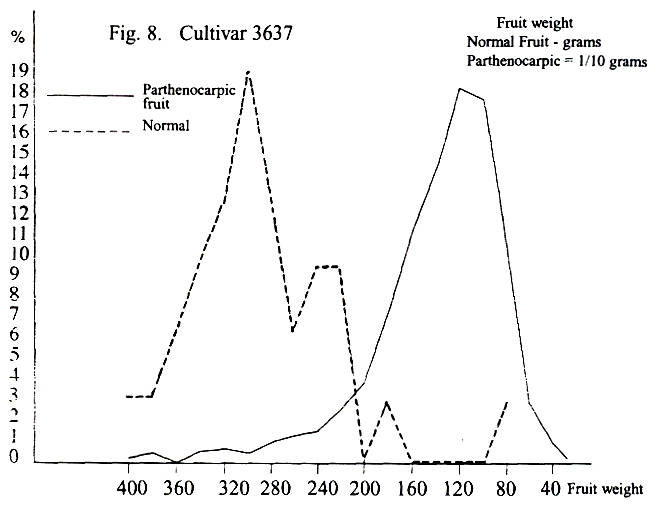
As noted previously, cultivar 3637 seems to be in a
category all by itself. This would appear true for both parthenocarpic
and for normal fruit. A knowledge of what the tree
underwent during the previous winter can maybe explain the reason for the
apparent difference in the distribution curve between 3637 and the remaining
cultivars. During the previous winter, water was standing around the tree on
three sides for a considerable period. This could have caused asphyxiation and
death of many roots. This would lead during the early part of flowering to an
insufficient moisture uptake and inhibition of fruit set. Thus, in the curve of
the normal fruit there is a possible sign of a peak at the 400 gram mark, a
rise to the maximum, and then three peaks as the fruit size drops. In the case
of the parthenocarpic fruit, there are two very small
peaks (at 38 and 32 grams) before the maximum, and then a steep drop to zero. Parthenocarpic fruit of less than two grams weight would
most likely be too weak to survive—the end result being that some of the peaks
have simply not developed at all.
The distribution curves can be also grouped thus:
3637 with few peaks, possibly with some which have not survived; at the other
end, the two cultivars which bore numerous normal fruit, with up to nine peaks;
and the remainder with five or six peaks. If we now examine the minimum
temperature regime, we find that for the total flowering period of the trees
examined (20th March-10th May), there are nine major
peaks of temperature. There also two very tiny peaks (17th and 25th April)
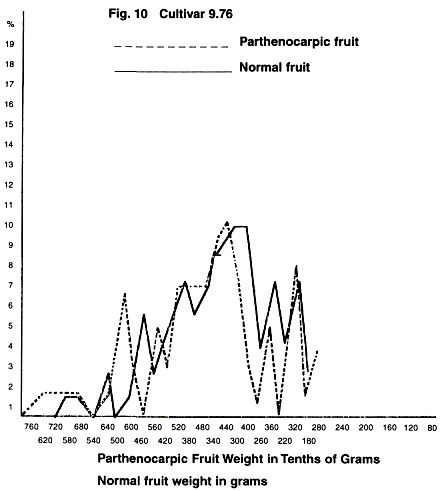
which it is felt can be ignored. In this way, we have the same number of
minimum temperature peaks as we have size distribution peaks in cultivars 9.76
and 125.06. The remaining cultivars had finished flowering by the 30th of
April, and therefore the last three minimum temperature peaks could not have
affected their fruit set, and thus they have fewer peaks. This situation is
what would be expected if minimum temperature13 and fruit set time14 affect
fruit size.
As will be shown in the part on normal fruit, the
number of peaks varies each year depending on the number of minimum temperature
peaks during the flowering period. As has been suggested with cultivar 3637,
one or more of the other three climatic factors controlling yield (heat stress,
March soil temperature, and winter rainfall—this latter could also be called
poor soil aeration7) can cause erasure of one or more of the distribution curve
peaks due to bringing about the drop of the weakest fruitlets.
Length of the flowering period will control the maximum possible peaks in the
curve.
Steyn et al. have found that E-type trees have
earlier flowering commencement (11). This author has noted that trees which
have borne low crops tend to commence flowering earlier than trees of the same
cultivar which have borne moderate to good crops, while in some cases trees
which have borne very heavy crops do not flower at all in the following spring.
A crop of parthenocarpic fruit could be considered as
acting as a low level crop as regards the nutritional balance of the tree.
Earlier and prolific flowering could give rise to in-tree competition which in
itself could lead to weaker fruit set. Some cultivars give numerous parthenocarpic fruit, while others do so only rarely. Steyn et al. (11)
point out the possibility that fruit development is controlled by phytohormones, and that possibly instead of their being
supplied by the developing seed, the tree itself or the mesocarp
of the young fruitlet can supply sufficient hormone
for the fruitlet to "hold on" and develop
into a seedless fruit. A cultivar which has a greater ability to supply the
necessary amount of hormone from outside the seed will be more likely to
produce a crop of parthenocarpic fruit than one which
does not have the hormone producing capability. That these phytohormones
(cytokinins and gibberellins) are present could be
inferred from the following: Martin et
al. (8) have found that these hormones can be used to cause elongation in
apple fruit. According to the work of Zilka and Klein
(14), later set Hass avocado fruits tend to be longer than early set fruit.
Field observations by this author have indicated also that later set fruit
tends to be longer and less round than early set fruit.
Whether too low minimum temperature causes a
cessation of the reproductive process or if the fruitlet
needs a certain length of time above a critical minimum temperature level in
order to complete certain essential processes before development can proceed remains to be clarified. However, what does seem to
be certain is that minimum temperature controls fruit set. The earlier a fruit
sets, the larger it will be able to become, and this is true of normal as well
as parthenocarpic fruit. Any condition, nutritional
in the tree or external in the soil or air, causing a disturbance of the
fragile reproductive process will reduce the "hanging on" power of
the young, newly fertilized flower-fruitlet; but the
minimum temperature level will control the number of flowers being productively
fertilized. It is suggested that parthenocarpic fruit
are those fruitlets which through meteorological
conditions were unable to pass through the critical stage in the early
development of the seed, but due to the hormone production of the tree managed
to "hang on."
The fact that cultivars flowering after the first of
May are in general good yielders (e.g.; 125.06 and
9.76) does not seem to concur with the distribution curves, for it would be
natural to expect that these cultivars would have the largest peak at the end,
or small fruit side, of the curve, and this was only found with the parthenocarpic fruit of 122.16 which finished flowering
early. Could it be that later flowering and better yields are genetically
linked?
Different parts of the tree have slightly different
temperature regimes, depending on tree side (toward or away from the sun),
height above ground, or climatic conditions. As a result, it is possible to
find on the same tree slightly different flowering regimes, not only as to when
a particular area of the tree begins to flower, but also the opening and
closing of the various stages (this is based partly on unpublished data of Lomas and Zamet). Thus, each
flower, depending on the cultivar, will have slightly different reaction to
cold or warm spells. Each flower opens at a slightly different time, and is
liable to be pollinated at a different time. Small differences such as these
could cause the difference between normal or parthenocarpic
fruit set, or simply total flower or fruitlet death.
One cannot ignore the fact that the actual size a
fruit will grow to depends not only on the cultivar and the temperature regime,
but also on its position on the tree, the number of fruit on the tree, the
number of fruit on a particular branch, and the number and size of leaves
supplying photosynthates to any particular fruit, and
finally to the light regime in the area of the said leaves. Nevertheless, with
the present total sample of over 2,000 fruits and seven cultivars, the evidence
for waves of fruit set seems very strong; and their similarity to the number of
minimum temperature peaks (with regard to the flowering period) is sufficient
to indicate a strong possibility that it is these minimum temperature peaks
which are the cause of the waves of fruit set. Avocado growers, at least in
Literature
Cited
1.
Anon. 1976. Research in progress.
2.
Babcock, E. B., R. E. Clausen. 1927. Genetics in Relation to Agriculture.
McGraw-Hill Book Company.
3. Blumenfeld, A., S. Gazit. 1974.
Development of seeded and seedless avocado fruits. J. Am. Soc Hort. Sci. 99:442-448.
4.
Ish-Am, G. 1985. Avocado Pollination by Honey Bees in the Fuerte, Ettinger and Hass
Varieties. Thesis submitted toward the M.Sc.
degree of
5.
Lahav E., D. N. Zamet. 1976. Flower, fruitlet, and fruit drop from avocado trees, pp. 57-63. In Research in Subtropical Fruit Trees. The
6. Lomas, J. 1988. An agrometeorological
model for assessing the effect of heat stress during flowering and early
fruit-set on avocado yields. J. Am. Soc. Hort.
Sci. 113(1):172-176.
7. Lomas, J., D. N. Zamet. 1994. Long
term analysis and modeling of agro-climatic effects on national avocado yields
in
8.
Martin G.C., D. S. Brown, M. M. Nelson. 1970. Apple shape changing possible
with cytokinin and gibberellin
sprays.
9.
Lovatt, C. 1994. Report on a research symposium in
10. Sedgley M., C. M. Annells. 1981.
Flowering and fruit-set response to temperature in the avocado cultivar 'Hass'.
Sci.
Hortis. 14:27-33.
11. Steyn, E. M. A., P. J. Robbertse, D. Smith.
1993. An anatomical study of ovary-to-cuke development in consistently low producing trees of the
'Fuerte' avocado (Persea
12. Tomer, E., S. Gazit, D.
Eisenstein. 1980. Seedless fruit in 'Fuerte' and 'Ettinger' avocado. J. Am. Soc. Hort.
Sci.105:341-346.
13. Zamet, D. N. 1990. The effect of minimum temperature on
avocado yields.
14. Zilka, S., L. Klein. 1987. Growth kinetics and
determination of shape and size of small and large avocado fruits of the
cultivar 'Hass' on the tree. Scientia Hort. 32:195-202.